����������:
1) ������������� ����������������� ������ ��������� �������� �� ������ ������� ������������ � ��������� �� ����� � ������������ ���� (���������� �����������, ������� - �.�. ���������);
2) ����� ����� �� ����� ��������������� �� ������������� ��������, ���������� � ������;
3) ������ ������ �������� ��������� � �� ����������� �������� ������ � ��������� ������� � �������� ������������ �� ���� www.ecosystema.ru
������ ������ ���� ������������ � ������� �������������� ������ ������������� �����
"������� ����"
��������� ��������� ����������
������: ����� �������������
�.�. �������, �.�. ��������
2003
������ ��� ���������� ���������
����������������� � ����� ��������
�������������. ���� ��������������� �����
������� ��������� ��� �� �������� ���������. �
��������� �������������� ������������
������������, ���� ����������� ������
������������� � ��������, ������� �����������
�������� �����.
������ ������� ����������������� � ����������
����� ����� �����������. �� ��������� ��������
������ ���������, ��������� ���� ����� ����� �
���������. ����� ����� ����������� ������
�������� ������ �� ������� ���������
�������������, �������� ��������� � ��������
���������.
�� ������ ����������� ���������� ������������
���������� � ������ ����� ����� � �����
����������� ���� ������������� �� ������
���������� �������������. � �������� ������ ���
������������� ���������� �� ���� ����������
�����.
���� ������ ������������ � �� �������
��������� ���������� ������ �������
�������������� ������ �������������: �����
�������� �������������, �� ������� ���
���������� ��� �����������, � ���������
���������� ������������� �������� ��
����������� �����������. ����������
������������ ������ �� ����� �������.
����� ������������� � ��� ���������
��������� ����������� ����, ������������� �
����� ��������������� �����������,
������������, � ����� �������, �������
����������� ������ ���������, � ������ �������,
� ����������� ��������� �������������,
��������������� � ������� ��������� � ���,
������� ����������� �������� � �����������
������������� ������������ (�����������,
���������, 1983).
��������� ����� ����� �������� ����������� �
����������� ����� ���������� � ����� ����, �
�������� 30-55 ���� (�������, 1991). ������,
��������������� ����� ������� �����������,
��������� ����� ����� �������� ���� ����� �
���������� ����� ������ ����������. ���� �������
������ ����� ����������� ��������� ������
������������ ������� ����� � ������
�������������� ������� �������� ������. ��
�������� � �������������� ������������
��������������� ��������� ��������� ���������.
����� �������, �����, �������� ����������
������, �������� ���������������� �����
�������� ��, �.�. �������� �������� ���� �� ������
����� �� �� ������������� ������ ������,
����������� �������. ������, ��������
����������� ����� ��������� ������� � ��������
����������, �� ������� ���������� � �������.
�����, ����������� �������, ���������� ��
��������� ��������� ��������.
�������� ���������� ���������� ����������� ��
���������� ����������, ����������� ���� �����.
���� �� ���� ������ ���� ����� ���������
���������������� ������� �� �����
�������������. �������� ��������� �����������
��� ����������� ����� ������, ��������,
��������� ������� �������� �������, ��������
����������� �� ������ ��������� �������� �����.
������������ ������� �������� �� ����� �����
����������� ��� ������ ��������� ����������. �
�������� ����� �������, �������� �� �����
�������������, �����������, � ���������, �������
��������� (�������, 1993).
�� ��������������� ������� ��������� �����
����������� ������ ������, ������� ��� �������
���������� ����� � ������� ����. �����,
����������� �������, ��������� ������ ��������
������� � ������������ ������ ������ ������.
����� ���� ������������� ������� �����������
����������, �������� �������� ��� ����������
������ �� ���� ���������� ���������� ������
����������. �� ������ ����� ���������� ���������
��� ������� ����� �� ������� ������ ����������,
��� ��� � ����� ������ ����� ����������� �����,
����������, ��� ���������� ������, ������������
�� �������� � �������� � ������ ������
���������� ������� (������, 1990).
�������������, ������������ �������
�������������, ���������� �������, ����� �������
����������� �������� ��� ������, ��� �������
������� �� �� ������ �� �������. �������� �
�������� ���������� � ���� ��������������
�������������� ����������� �������������
�����������. ������ � ���, �������������
������������� ����������� ������ �� ����������
�����������, ��� �������� � ����� �������
���������� ����������� � �������������
��������������.
������ ������������
�� ������� ����� ������������� �
���������������� ��������� ���������� ������
(Turdus sibiricus Pall.).
����� ���������� ������ � ������ � ����������
�������� �������. �� ������� ������ ��� ����� �
�������������� ���������� � �����������
��������� ��������� ��������. � ����������
�������� ����������� � ������ ��������� ���, ��
�������� ����������� �� �������� ����� ������,
��� ��������-������������ ��������� ����������
� ������ ������ ���������� � ��������.
������� ���� ������� �� ����� ����������� �
������ �������� ������ � 31 ���. ��� ���������
�����, ��� ��� ������ �������. ������� �����
���������� � ����������� ������� �� ���������
������� �������� ���������. ��������� ��
����������� ��������� ������, ��� � �����������
�������������� ��������� 1 ���� �� ������. �����
������ ����������� ���������� ������ ����������
�� ��������� ��������� ���������. � ��������
���� ����� ��������� �������, �������� ��������
� ��������, ������������ �������������. ����� �
����� ��������� ������ ������� � ����������� �
������������� �������, �������� �������.
����� ���������� ������� ����������
�������������� ����������, �� ����� ��������
������� ����� �������� ��������� �������, ��
������� ������������ � ��������� ����.
��������������� ����� ������ �� ����� �������
� ������ ������ ������� � ������ ������
��������. ������ ����� �������� �����������
�����, ��������� � �������� ������� (��������� �
��., 1954; �������� � ��., 1989; Clement, Hathway, 2000).
���������� ������
����� �������� ������������� � ������
���������� ������������� ��������� �������, ��
������� �������� ������������� �� ���������
���������. ��� ������ ������� ���������
�����������, ��� ������ ����� �������������
������������� ����, ������� ��������� �������,
�� ����, ������������ ��� �����������. �� ������
������������� �� ������� �� ����, � ���������
����� �������� ���������� � ���� ����������
��������. ����� �������, ������� �����������
��������� ����������� �������� ���������
�������� ��������� �������������.
���� ��������� �������� �������������, ��
������ ���������, ��������� ��� ��������
���������. � ��������� �������������� ��
��������� ��������� �� ������������� ������� ��
���� ���������.
1. ���� ������ ������. ������������� ����
������� ����������: ��� ������, ��� ������
��������, ������ ���������� � ������ ������� ���
�������� ������� �������.
2. �������� ������. ��� ������ ���, ��� ����
�������������� ����������� ����� � �����������
��������� ������ �������.
3. �������� ����� �������. ��� ������ ����� � ���
����� ��� �����, ��� ��� ������.
4. ����� ���������� �������. ��� ������ ��������
� ��� ����� �������� ������, ��� ������
����������� ��������� � ������ ������������
����� ���������.
5. ������� ���������. ������� ������������
����� ������������ �� ������ �������. �����,
������������ ��������, ���� ������ �����
�����������. ������������ ������, ������� ����
������� � ������� �������� ����������.
�������� ���� ����������� �����������
��������� �� ������, ������������� �� �������
��������� �������� ������� ��������
�������������. �� ������������, ��� ��� �����
����� ���� �������� �� ��������� ��������.
1. � ������ �������������� �������������
������� �����: ����� ������, �������, �������.
��� �������� �������� ������� ���������
���������� � �� ��������� ���������� ���������.
2. ����� ����� �������� ������������������
�������������, �� ����� ����������� � ��� ����
��-�� �����������.
3. � ������ ������������� ������� �����������
��������� ����, �� � ��������� ���� ������� �����
������������ �����.
4. � ���� � ������ ������������ ����������� ���
����� ����������� ������� ������ �������������,
� �������� ����������� �� �����������
�������������.
��������� � ������
�����, �����. �������� ������ �� ����������
������������� ������� ��������� �������
�������� � ��������, � ������������ �. ������
������������ ������ ������������� ���� (62�20 �.�.,
89�00 �.�.). � ������ ������������ ��� �����������
������ �� 2002 �., ��� � ��������, ��������� �.�.
�������, �. �����������, �.�. �������� � ������� �
1991-2002 ��.
���� ������
�������� �������������. �� �������
��������, � �������� ������� �� ���������
��������� �������, ���� ������� ���������������
��������, ������� �������� � ���� ���������
���������� (�������� ������, 1998 �.).
��������������� �������� �������� ��������
������ ����������� (������������ ��������) �
������ �� ���� �������� ������ ��������� ���
������� �� ������������� �����. ����� ����,
����������� ���������� ��������� ��� �������
���� �� �����.
�������� ��������� �����������. ���
�������� ������ ������������� � ����������
������ �� �������� ����������� ���������
����������� � �������� ������������� �� �������
� 50 ��, ��� � ������� 12 ��� ����������� ���������
������������ ��������� ���������� (��������
������).
���������� �� ��������. �� ��� ��
������� � 50 �� � ����� ������ �������� 3
�������������. � 2002 �. �� ������� ����������
����������� ���������� ������ � ����� ���������,
��������� �������������� ��������
������������� ������������� ������������
�����������.
�� ������ ����������� ��� � ������
������������� ������� ������� 21 ������. ������
����������� ������ ������ 1-2 ���. ���������
���������� ������. ���������� ������� ������ �
��������� �� 0,1 �.
�������� ���������� ���������: ���� �� ��� ����
������� �����, ��� ������ ����������� ����������
�� �������. �� �������� �� ������ ��������� ��
��� ���������: �����-���������� � ������ �����,
����������� ��������.
��������� ������
�������� �������������. ��� ��������
������������� ������� ��������� �������������
�� �������� ��������� ����������. ��� ���������
��������� � �������� ������ ������� �����
��������. ������ ������ ������ ��������
���������, ���������� ����������� �� ������
�����. � ���������� ������ ������� ����
������������ ������� �� ������� ������� (�������
�������� �������� 50�50 �). ��� ���������
���������� � ���������� ��������� �����, �
���������������� ������� ���� � ���������
������.
��������� �����������. ���������
��������� �������� �������� ����� ����������
������ �� 12-������ ������ � ���������� �
����������� ���/�� ��� ������� �������� ��������
�������� 50�50 �. ������� ��������� ����������
��������� ������� ������ ��������, � ����� �
������� ���������� � ������ �������� ����������
������� ��������. ������� ������ ���������
������������� � ��������� ��������� ���������
��� ������� �������� ��� 9 ������� ���������.
����� ������� ���� �������� 180 ��������
��������� � 180 �������� ������ ��� �������
��������� �������� ��������������. ��
����������� ���� �� ����� ��������� �������
��������������� � �������������� �������.
��������� ����������. ��� ��������
�������� ����� �������, �������� ���������
���������� ��������� � �����������, ����������
������ ������ ������� ������� �� ������ �������
�����������. ��� ����������� ������� ��������
������� ��������� ���������, ���������
�������������� ���������� ������ ������
���������� ������ � ����������� ���������, �
����������� ��������� ������������ �������. ���
���� ����� ������ ��������� ��������, ��
���������� ������� � �������� ������� �������,
�� ���� ���, ��� ������� �������� �����
���������� � ������� �� ��������� � �������� � ��
������ �� ������ �� ������. �� ������ �� �� ���� �
�������� �� ��������� ������ ����������� �����
�� ��������. ��������� ��������, ��� ���� �������
� ���� ����� ����������� �� ���������� �� �
������, �� �������, ��� ����� ����������� �����
���� �� �������� ������������� S-�������� ������
������������� ����� (������ ��������������
�����) (����, 1975; �����, 1980):
M = K / (t + e a-rt) , ��� M � ����� ����, t � ������� �
���� �� ������� ����������, � ��������� ����� �
���������, ������� ������� ���������� ��
����������� ������. ����� ��� K � �������������
����� ����, a � ����������� ����� ��� ����������,
r � ����������� �����.
������������ ���� ���������� ����������
���������� ������ � ��������� STATISTICA. ����
�������� ������� �� ������� ������������� �����
�������, ����� ����� ��������� ���������� ��
����� �� ������ ���� ����������. ���� ��������
���� �������� ��������� ����������� ����� (���.
1). ���������� � ���� �������� ����� ������ ���
������ �����������, �� ���������� ��� �������
������, ��� ��� ����� ������� �� ����.
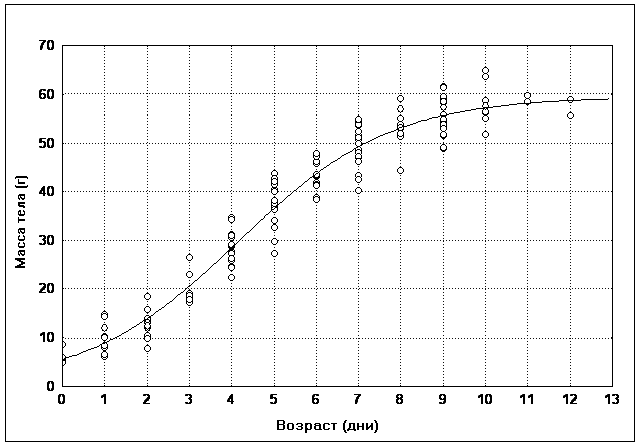
��� 1. ��������� ���� �������, ��
������� ������� �������� ������ ������
�� ��������� ����� �� ���� ������ �����������
������� (����� �������, ��� ��������������� ���
����������� ��������) ���������� ������ �����
������� � ��������� ��������� (���. 2).
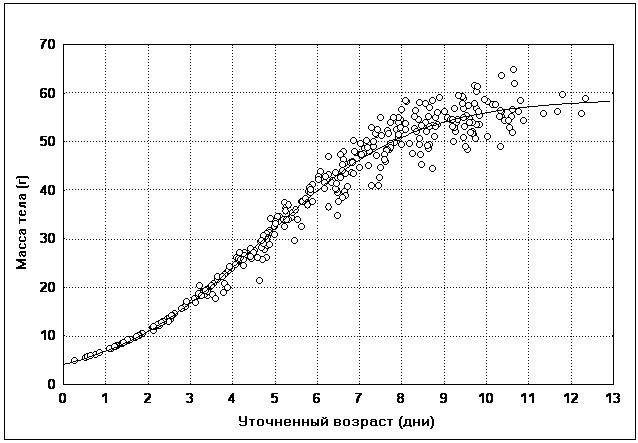
��� 2. ��������� ���� ������� �
���������
��� ������ ������������ ������� ���������� ��
�������� � ���� �� ������ ����. �������� �������
� ���� � ������ �������� ����������, � ��������
��������� ���������, � � ����� ����� �����������,
��������� ������ ������� ���� ������ ��
����������� �����, �������� �������� � ������
������� (���. 3).
��� 3. ���������� �������� ������� �
����
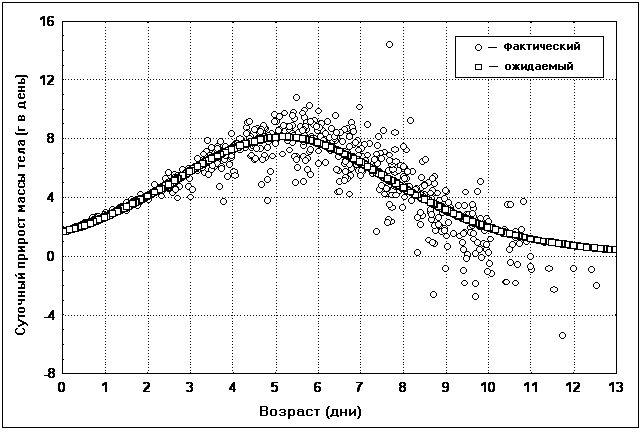
������� ��� ��������� ������������ ������� ��
������ ������ �������� �� ������������
���������� ������������ �������� � ��������� ��
���������� �������� (���. 4). �����
��������������� ���������� �������� ������ �
����� �� ������ �������������, ������������
������� ������������� ������� �� ����� �
�������� �� 1 �� 10 ����. ��������� � ��������
������� ����� � ������ ��������� ��-�� �������
������������� �� ������ ���� ��������, � �����
��-�� ����, ��� �������� ��������� �������
������� �������, ����� ������������� �� �����.
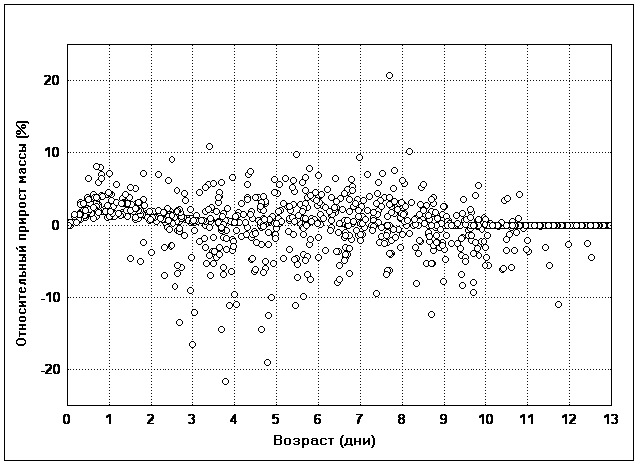
��� 4. ���������� �� ������� �������� �
���� ��� ������� ������
����������
����������� ��������� ��������� �� ���������
�������������
���������� ������ ������� ��� ����������, ���
��������� ��������� ���������� ������ �
��������� ������ ������������ ������������ (���
. 5).
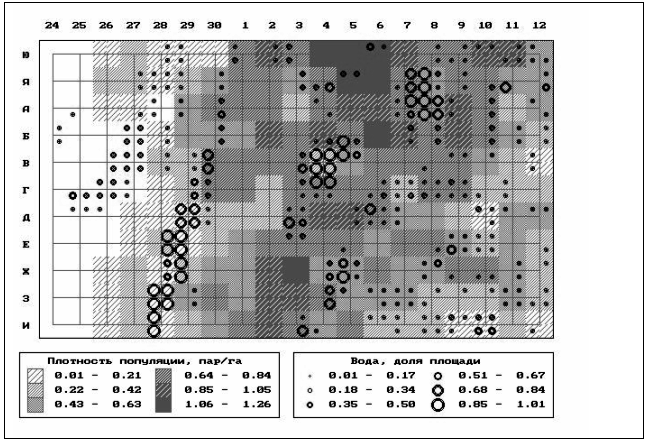
��� 5. ������� ����������� ���������
��������� ���������� ������ �� ���������
�������.
������������ ����� ������������� �������� ��
�������� 50 � 50 � �� ���������,
� ������� ��������� �������� ������������� �
����� ����.
��� ���������� ���������� �� ������ ������,
��������� �������� ������������ ���������
(����� 24-30 �� ���. 5), �������� 0,7 ���/�� � ������,
������������ ������� ������ ����� (����� 1-2).
����� ������� ��������� (�� 1,0 ���/��) �������� �
����������� �����-�������� ���� � ������
��������� �� �������� � ���������� �����
������������ ���� (����� 3-5) � ������ ��������
����� � �����. ����� ������ ����� ���������
���������, �� ����� �������. ����� �������������
������������ ����� ������������� ������������
�����-��������� ������ � ������ ��������� ��
���������� (����� 6-12). ��� ������, � ����� ��
��������� �����, ��������� ����� �� ���������.
�� �������� ������������� ���������
����������� ���������� ������ � ���������
������������� �� ���� ������� ���������
��������. ���������� ��������� � ����
������������� ���������� �������� � ����. 1.
������� 1. ���������� �������
����������� ��������� ��������� ����������
������ � ���������� �������������� �� 180 �������
���������
(r � ����������� �������� ���������� �������, R2
� ������� ����������)
�������� ������������� |
r |
R2 |
| ���������� ��� �������� ������� ���� |
0,86 |
0,74 |
| ������������� ������ ������ |
0,66 |
0,43 |
| ������������ ������ |
0,64 |
0,41 |
| ����������� |
0,11 |
0,01 |
| ����� ����� �������������� |
0,57 |
0,33 |
| �������� � �������� |
0,75 |
0,57 |
| 1 ���� ��������� (12-25 �) |
0,33 |
0,11 |
| 2 ���� ��������� (5-12 �) |
0,45 |
0,2 |
| 3 ����: ������� ���������� (2-5 �) |
0,64 |
0,41 |
| 4 ����: ������ ���������� (1-2 �) |
0,62 |
0,38 |
| 5 ����: �������� ������ |
-0,55 |
0,31 |
| ������� (��� � �����) |
0,38 |
0,15 |
| ������ |
0,21 |
0,04 |
| ����������� ���������� (�����, ��������� �
��������) |
0,67 |
0,45 |
| ������������ ���������� (���� ��) |
-0,33 |
0,11 |
| �������� |
0,75 |
0,56 |
| ����� |
-0,64 |
0,41 |
| ��������� |
0,47 |
0,22 |
| �������� �������� ������ |
0,82 |
0,67 |
��� ����� �� �������, ��������� ����������� ��
������� ���������. ����� ������� �����
�������������� � ������� ��� ������� ����. �� �
������� ��� ������� ��������� �����������
��������. ��������� �������� �������� ������
��������� � ��������� �������������� �� ������ �
������� ������ ������������� ���������. �� �����
����� �� ����. 2.
������� 2. ���������� ������� ���������
�������� �� ��������� ���������. b �
������������ ��������� ���������, b � �����
��������� ��������, r � �� ������� ��� ������
������ ��������, R2 � ������� ����������
| |
������ ������� |
b |
b |
r |
R2 |
������� ���������
�������������� |
| ��������� ���� |
-0,79 |
|
|
69,50% |
| 1 ���� ��������� (12-25 �) |
-0,4 |
-0,65 |
-0,55 |
| 2 ���� ��������� (5-12 �) |
0,27 |
0,34 |
0,41 |
| 3 ����: ������� ���������� (2-5 �) |
0,43 |
0,2 |
0,23 |
| 4 ����: ������ ���������� (1-2 �) |
1,99 |
0,85 |
0,67 |
| 5 ����: �������� ������ |
-0,03 |
-0,21 |
-0,23 |
����� ��������� ������������� |
| ��������� ���� |
-0,19 |
|
|
76,90% |
| ���������� ��� �������� ������� ���� |
0,25 |
1,12 |
0,78 |
| ������������ ������ |
-0,77 |
-0,31 |
-0,33 |
������� ������ ��������������
|
| ��������� ���� |
0,49 |
|
|
78,10% |
| ������� (��� � �����) |
-0,49 |
-0,55 |
-0,45 |
| ����������� ���������� (�����, ��������� �
��������) |
0,53 |
1,16 |
0,68 |
| ������������ ���������� (���� ��) |
0,12 |
0,22 |
0,25 |
| ����� |
-17,16 |
-0,28 |
-0,43 |
| ��������� |
4,28 |
0,24 |
0,41 |
�������� ��������� ��������� |
| ��������� ���� |
-0,75 |
|
|
87,50% |
| �������� �������� ������ |
1,9 |
0,82 |
0,78 |
| ����������� ���������� (�����, ��������� �
��������) |
0,36 |
0,79 |
0,65 |
| 1 ���� ��������� (12-25 �) |
-0,4 |
-0,66 |
-0,7 |
| ������������ ���������� (���� ��) |
0,19 |
0,32 |
0,48 |
������� ��������� �������������� ���������
��������� ��������� ��������� ����� �� 70%.
�������� ����������� ������� ����������� �
������������� �������� ���������� �����.
������������� �� �� ������� �������
����������������� �� ���. 6 � 7.
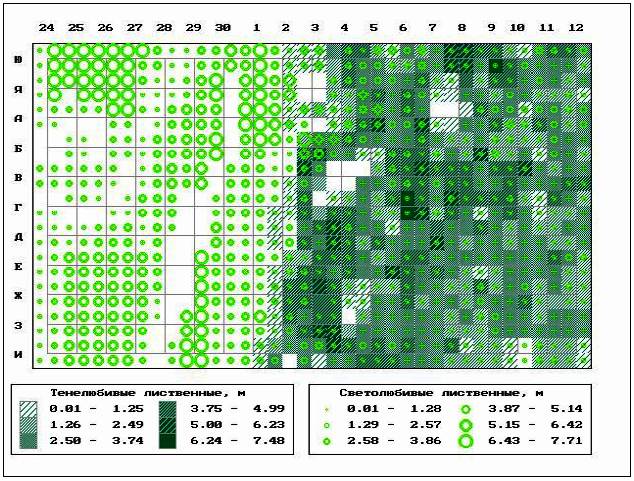
��� 6. ���������� ����������� (�����,
��������� � ��������) � ������������ (���� ��)
���������� ����� �� ��������� �������.
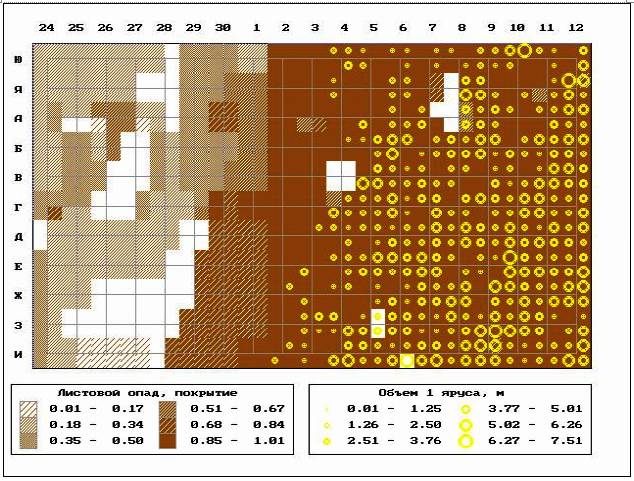
��� 7. �������� �������� ������ �
���������� ������� ����� ��������� (12-25 �) ��
��������� �������.
��������� ���������, ������������ ��������
���� ������� ��������� �������������, ���������
����������� 87,5% ��������� ������� ���������
���������� ������ � ������ ����������� (���. 8).
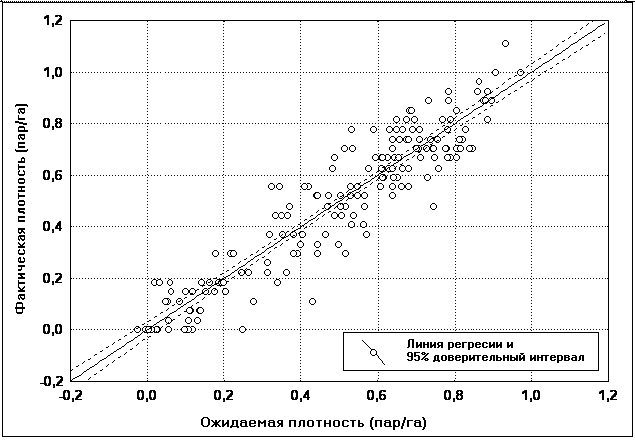
��� 8. ������������ ��������� ���������
�� ������ ��������� ���������.
������� 3. �������������� ��������
������� ������������� ������ ��������� �����
������ (�����) |
������������� |
�������� ������ |
������������� �������� ����� |
����� ������� |
������� |
> |
+ |
29� |
1 |
5 |
� |
0 |
? |
1 |
29� |
1 |
4 |
-0,54 |
2 |
? |
2 |
29� |
1 |
� |
� |
0 |
1 |
1 |
1� |
1 |
4 |
0,86 |
2 |
2 |
2 |
1� |
1 |
5 |
0,52 |
3 |
1 |
1 |
1�0 |
1 |
5 |
� |
0 |
1 |
1 |
1� |
1 |
5 |
-1,58 |
4 |
2 |
1 |
2� |
1 |
5 |
-1,1 |
4 |
2 |
? |
2� |
1 |
5 |
-0,79 |
5 |
2 |
2 |
3� |
2 |
5 |
1,29 |
0 |
1 |
2 |
4� |
2 |
5 |
-0,44 |
4 |
2 |
? |
4� |
2 |
3 |
-0,91 |
1 |
1 |
2 |
4� |
2 |
5 |
-0,5 |
4 |
2 |
2 |
5� |
2 |
5 |
0,44 |
5 |
? |
1 |
5� |
2 |
5 |
0,54 |
2 |
2 |
1 |
5� |
2 |
5 |
0,94 |
4 |
? |
1 |
6� |
2 |
5 |
� |
0 |
1 |
1 |
7� |
2 |
5 |
-0,28 |
3 |
1 |
1 |
7� |
3 |
6 |
0,9 |
5 |
2 |
? |
7� |
3 |
4 |
1,59 |
4 |
2 |
? |
9� |
3 |
5 |
-0,32 |
1 |
2 |
? |
10� |
3 |
5 |
0,9 |
5 |
? |
2 |
11� |
3 |
5 |
0,01 |
5 |
2 |
? |
11� |
3 |
5 |
0,63 |
5 |
2 |
? |
12� |
3 |
5 |
0,57 |
4 |
1 |
? |
13� |
3 |
5 |
� |
0 |
? |
? |
����������� ���������� ����������� ��
�������� �������������
������������ ������� �������� � 2002 �. ���������
��� ���������� �� ��� ���������� 29 �����������
��� ��������� �������. � 24 �� ��� ����������
������, ������ � ���� ���, ������ ������� ����
�������� � ����� ������ �����������, ������� �
������ ������ (���. 9).
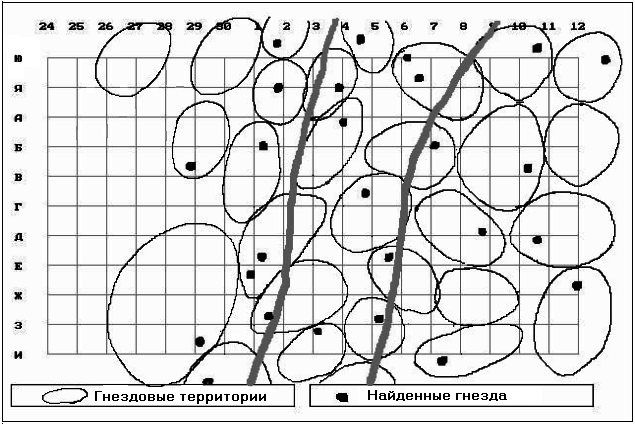
��� 9. ������������ ���������
���������� � ��������� ����� � �����������
��������� ���������� ������ � 2002 �.
������� ����� ���������� ������� �������������
(����� �������: 1 � ������ �����, 2 � �����������
��� � 3 � ������� �����).
������������ ������������� ������������
��������� ������������� �� �������� (���. 6 � 7), ��
�����, ��� �� ��������� �������� ��� ��������
���� �������������.
������������� 1 (������ �����) ���������� ��
���� ������ ����������� �������� ������� �����,
������������� ��� ����� �������� � �����������,
���������� �������� ������.
������������� 3 ���������� ��������� ������
������� � ������� ��������� ����� ����������.
��� ����������� � ������� ����������, ���� �����
������� ������� ����� ��������� �����.
� �������, ���������� � ���������
��������������, �������� ��������� �����������
����������� (����. 4).
������� 4. �������������� ������������
� ����������� ����������� � ��������� �������
| |
������ ������� |
b |
b |
r |
R2 |
������� ���������
�������������� |
| ��������� ���� |
-0,79 |
|
|
69,50% |
| 1 ���� ��������� (12-25 �) |
-0,4 |
-0,65 |
-0,55 |
| 2 ���� ��������� (5-12 �) |
0,27 |
0,34 |
0,41 |
| 3 ����: ������� ���������� (2-5 �) |
0,43 |
0,2 |
0,23 |
| 4 ����: ������ ���������� (1-2 �) |
1,99 |
0,85 |
0,67 |
| 5 ����: �������� ������ |
-0,03 |
-0,21 |
-0,23 |
����� ��������� ������������� |
| ��������� ���� |
-0,19 |
|
|
76,90% |
| ���������� ��� �������� ������� ���� |
0,25 |
1,12 |
0,78 |
| ������������ ������ |
-0,77 |
-0,31 |
-0,33 |
������� ������ ��������������
|
| ��������� ���� |
0,49 |
|
|
78,10% |
| ������� (��� � �����) |
-0,49 |
-0,55 |
-0,45 |
| ����������� ���������� (�����, ��������� �
��������) |
0,53 |
1,16 |
0,68 |
| ������������ ���������� (���� ��) |
0,12 |
0,22 |
0,25 |
| ����� |
-17,16 |
-0,28 |
-0,43 |
| ��������� |
4,28 |
0,24 |
0,41 |
�������� ��������� ��������� |
| ��������� ���� |
-0,75 |
|
|
87,50% |
| �������� �������� ������ |
1,9 |
0,82 |
0,78 |
| ����������� ���������� (�����, ��������� �
��������) |
0,36 |
0,79 |
0,65 |
| 1 ���� ��������� (12-25 �) |
-0,4 |
-0,66 |
-0,7 |
| ������������ ���������� (���� ��) |
0,19 |
0,32 |
0,48 |
�������� ���������� ����������� ��
��������������, �� �������� ���������
���������� (����. 5).
������� 5. ���������� ����������
����������� � ���� �������� ����� �������������
������������� ��������� � 2002 �. (�� ������ 26
�����).
���������� �������� ����� ����� ����� ���������
�� ������� ���������� ����������� � ���������
����� ������������� �� �������� t ���������.
������ ����������: - p > 0,1; + p < 0,1; * p < 0,05; ** p <
0,01; *** p < 0,001
| |
������������� (�����
����������) |
���������� �������� |
1. ������ ����� |
2. ����������� ��� |
3. ������� ����� |
1:02 |
2:03 |
3:01 |
| ���� ������ ������ (%) |
47% (15) |
40% (15) |
86% (7) |
- |
* |
+ |
| ������ ������ (��� ����) |
16,1 (9) |
13,3 (9) |
13,5 (8) |
- |
- |
- |
| �������� ������ (���) |
4,75 (8) |
4,78 (9) |
5,00 (8) |
- |
- |
- |
| ����� ������ (�������) |
2,22 (9) |
2,56 (9) |
3,63 (8) |
- |
- |
- |
| ������������� �������� ����� (%) |
-0,44 (6) |
0,13 (8) |
0,61 (7) |
- |
- |
* |
| ������� ��������� (������ �� 1 ��) |
0,49 (9) |
0,80 (9) |
0,59 (8) |
*** |
* |
- |
�� ���������� ����������� �������, ��� �
������� ����� ��������� ���������� ������
������ ������, ��� � ������ ����� ��� ��
����������� ����. ������ ������ � ������ �����
�������� ���������� � ������� �� 3 ��� ������, ���
� ������ �����, ���� ����� ������ ������������,
����� �������� ����������� ��������. �������
�������� ������ � ������������� 3-5 ���, ���
������� (�������� �� �������� ����) ������, ��� �
1-� � 2-� ��������������. ����� ������� ����������
� ������ �����, ������������� ������ ��
����������� ���� � �������� �� ������ ������
������ � ������� �����. �������� �� ���������
���� ����������� ���������� ����������� ��
����������. ������������� �������� �����
������� ���������� ������ � ������� �����, ��� �
������, �� �������� ����� 1-� � 2-�, 2-� � 3-�
��������� �� �������. ��������� ���������
�������� �� ����������� ���� ���������� ����,
��� � ������ � ������� �����, ������ ��������
����� 2-� � 3-� �������������� ����������� ���.
����������
���������� ���������� ��������� �� ���������
������������� ��������� �� �������� ���
����������� ���������� ������ � �������������
��������� ��������� �� ������ ������� ��������
����� ������. �� ����������, ��� ����������
��������� ��������� �������������, �� �������
����� � ��� ��� ���� ���� ������ �� �� ���������
�������� ��� �����������.
��� �������� ������� ���� � ������. ���, ������
�� ��� ����������� ���������� ��� ����������
������ ��������� � �������� �� ����� ���� (����.
5). �� �����������, ��� � ��������� ��������
��������� ���������� � ���� �����������. ������,
����� ��������������� ������� ���� �� ���� ���
�� ��������� ������ ���������.
������� �� ����������� ����� ����� ��������
���������, ������� ����� ���� ������ ������ �
��������� ������ �������������. ��� ����������
������, ����������� �������������� � ������
���������, ������� �������� ����� �������� ����
����������� ������� � ��������� � ��� ������
��������������. ���� ����������� ������������
��������, �������� �������� ������ � ��������
���������� ������� ��� ��������� �����.
�������� ����� ������� ��������� �� �������
����������� � ���, ��������, ���������� �
������������ ������� � ��� �����. ���������
��������� ���������� ���������������
����������� ���� ���� ����� � ����������� ��
���� �������� ���������� ������ ������� �����.
����������� ���� ����� ������ �����������
�������� �������������� ���������� � �������
������ �����.
��������� ����� �������� ������������ ���
��������� ����������. ����� ����, ������������
������������ �������� ������� ��������� �����.
��� ������ ����� ������� ������������ � ������
����������� �����, �, �������������, ������
�������� ���� ����� �����������. ������� ������
������������ ��� ����� ����������������
���������� ��� ��������� � ��������.
������ � ��� ��� ���������� ������ �����������
�������� �������� �������������. ���� ���������
�������� ���������, ��� ������� ����� ��������
����������� ��� �������� �������� ������� �
������������� � � ����� ������ ������� ���
������ �����. ��������� ����� ������� �������
����� ��� ���������� ������.
���� ���������� ��������� 3 ����������
������������� �� ������� �� ������������, ��
���������� ��������� ���������� �� �����������
����. �� �� ���������� ����������� ������
������������� ������������. ����� ��������, ���
�������� ������ � ���������� ������� ����������,
��� ������ ������ � ��� ������ �����, ������� �
����������� ���, � ������ � ������� �����. ������
���������� �� ���� ��������� �������
������������, ������ ��� ����� ������� ��������
����� ��������� �� ��� �� ����������.
����������� �� ������ �� �������������
�������� ����� ������� � ���� ������ ������ �
������ ��������������, ����� �������, ���
�������� ��������� �������� �� ����������� ����
����, ��� � ������ �����, � � ������� ����� ��� ���
����. ����� ����� ������������, ��� ��������
�����, ��������� �����, ����� ��������� ��������
� �������� ������������� � ������� �������
�����������, ���� ���� ��� ������� ���� � ������
����������.
������� 6. ����� ��������� ���������
������������� � �� ���������� ������������
����� ����
| |
���������� |
����������� |
| ������������� ������ ������ |
0,76 |
0,39 |
| ������������ ������ |
0,84 |
0,89 |
| ����������� |
0,24 |
0,13 |
| ����� ����� �������������� |
0,72 |
0,84 |
| �������� � �������� |
0,92 |
0,88 |
| 1 ���� ��������� (12-25 �) |
0,61 |
0,89 |
| 2 ���� ��������� (5-12 �) |
0,54 |
0,55 |
| 3 ����: ������� ���������� (2-5 �) |
0,63 |
0,56 |
| 4 ����: ������ ���������� (1-2 �) |
0,77 |
0,8 |
| 5 ����: �������� ������ |
-0,61 |
-0,51 |
| ������� (��� � �����) |
0,66 |
0,86 |
| ������ |
0,49 |
0,85 |
| ����������� ���������� (�����, ��������� �
��������) |
0,85 |
0,86 |
| ������������ ���������� (���� ��) |
-0,57 |
-0,69 |
| �������� |
0,83 |
0,51 |
| ����� |
-0,65 |
-0,65 |
| ��������� |
0,36 |
0,04 |
| �������� �������� ������ |
0,94 |
0,87 |
���� ������ � �������� ������������� ��
��������� ��������� �������� � ������� ����
������ ������, �� ������ ����������� ������
�����, ������� � ������� �����, � ������ �
����������� ���. ������ �����, ��� ��� ��������
�������� ������� �� ������� ������������
������������� ��-��� ��������� ���������.
�������� �������� ����, ������, ��������
����������� ����� ��������� � ������� ������
������� � �������� ���� ������������ ���������
�������. ���� �� ���������� ���� ������ �
������������� ������������� ������ ����, ���
������� � ������� �����, � ���� ������� ��� ��
�������������� �� ����. � �� �� ����� �������
����� ���������, ����� ������ ����� � ��������
������ ������ �������� ���������. ��� � ��������
� ���������� � ������ �����, � ����� �� ��������
������������ ����, ��������� ����� ���������
������ � �������.
����� �������, ��������� ���� ���� ���������
���������� ����������� � ��������� ���������
������������� ��������, ��� ������������ �����
���� ��������. �������� �� ��, ��� ���������
����������� �� ����������� ���� �����������
����, ��� � ������ � ������� �����, ������� �����
������� ������ �� � ������� ��������
������������ ���� ��� �������, � � ���, ��������,
��� ����������� ��� ������ ������������� ��
����. ������ �������������� ������� �������,
���������� �� ������� ��������� ���������
��������, ������������� �������� �������
������������ ���������� �����������. �� �����
����� ���������, ��� ����������� ��� ��������
������� �� �������� ��������� ����� ����
��������������� ��������. ����� �������
������������� �����, ���������� ������ ������.
����������
- ��������� �.�., ������� �.�., ������������ �.�.,
����������� �.�., ������������� �.�., �������� �.�.,
�������� �.�., ����������� �.�., ����������� �.�.,
������� �.�. 1954. ����� ���������� �����. ��� VI. ���
���. �.�. ���������� � �.�. ��������. � ���������
�����, �.: 792 �.
- �������� �.�., ������������� �.�., ������� �.�.,
����� �.�., ������� �.�. 1991. �����
�������������������� ����������� �����������. 2.
���������� �����. � ������������� ������� �
��������� ���������� �����. ����� �� ����, �.: 32-164.
- ������� �.�. 1993. ��������������� ��������� �
�������� ��������� ���� � ����������. � �����,
������������ : 296 �.
- ������ �.�. ������ �����. ���������� �������. �
������ ��������������, �.: 399 �.
- ������� �.�. 1991. ������ ���������� �����
������������ �����. � �����, �.: 176 �.
- Clement P., Hathway R. 2000. Thrushes. � Christopher Helm, A. & C. Black, London:
463 p.
***
������ ������ ���� ������������ � ������� �������������� ������ ������������� �����
"������� ����"
������ ����������������� ������, ��������� � ������������� ������ ����, ����� ����������
� ������� ����������������� ������. ������ ������� ����������������� ������ (����������������� �������), ����������� �� ����� �����, ����� ����������
���.
 ������� ������
"�������� ����"
��������� "Parus"
������ ���������
������� ������
"�������� ����"
��������� "Parus"
������ ���������
|
���� ��������� ������� � ������������ �� ����������� � ������ ������
����� ���������� � ����� ��������-�������� ��� ������� �
 ������������ �������� (��� PC-Windows) ������������ ���� ������,
���������� �������� � ����������� 206 ����� ���� ������� ������ (������� ����, �������, ������, ���� � ������), � ����� ������������ ��������� ����������� ����������� � ������� ����.
������������ �������� (��� PC-Windows) ������������ ���� ������,
���������� �������� � ����������� 206 ����� ���� ������� ������ (������� ����, �������, ������, ���� � ������), � ����� ������������ ��������� ����������� ����������� � ������� ����.
���������� ��� ���������� � ��������� Android:
����� ������� ������,
����� �� ���� ������ �
�������,
������ ���� ������ �
����� ������,
������ ���� ������ �
����� ������
(�� ����� ������� �� ��������� ���������� �� ������� �� ��������� ��������),
���������� ��� iPhone � iPad:
����� ������,
������������ ���� ������ (Birds of Europe),
������ ���� ������,
����� �� ���� ������
� ���� �������� �������,
������������ ������� ���� �
������������ ���� �� ����
(��� �� ����� ,
��������� ������� ������������
����� ������� ������ (160 �����),
����� ������ (278 �����),
������� ��������������� �������
���������� �����,
�������� �����,
�����-������������ ����� "������������ ������� ������":
�����,
�������� ��������,
MP3-���� � �������� ���� (�����, �����, �������):
������ ���� ������� ������ ������ (343 ���� ����).
|
|

