����������:
1) ������������� ����������������� ������ ��������� �������� �� ������ ������� ������������ � ��������� �� ����� � ������������ ���� (���������� �����������, ������� - �.�. ���������);
2) ����� ����� �� ����� ��������������� �� ������������� ��������, ���������� � ������;
3) ������ ������ �������� ��������� � �� ����������� �������� ������ � ��������� ������� � �������� ������������ �� ���� www.ecosystema.ru
������ ������ ���� ������������ � ������� �������������� ������ ������������� �����
"������� ����"
���������� ����� � �������������
�������: ������� ���������� ����
� ����� ������ ������������ �����������
�������������� 1
�.�. �������
(������������ �������� ���, �.-���������)
2000
����� ������� � ������� ���������� �����
� ��� ��� ��� �.�.�������� (1931) �������������
������� � ���������� (�������������) ����� (��)
� ���������� �������������� ������, �������
����� ���� �������� �� ����� ������ ������� ����
�� ������ ��������������� ��������� � ����� ��
���������������� ����� �������������
���������� �������� ������������ ���� ���
������������ ��������� ������. �...������������
�����, �� (��) ���������, �������������� �� ����
���������� ����������� ������ ������, �������
���� ��������� ������������ ����������� �
����������� �� ����� ������� ������������.
�������� � ����������� ��������� � ��������
������ ������������� ���������� ����������
�����... ������ ������������ ����������
��������������� � �������� ������ ����������
����� ������������� ������������ ������
�������� �������... �������������� ��������
����� ���������� ��������� ������ ������������
���������������� ��������� ������������
������� ������� �������������...� (��������, 1974,
�.185-186).
����������� ����������� ������ ������ �
����������� �������� �������� ����� ��
(��������� ���������� �������������� ��������
�� ���������� ��������), ������ ��� ���� ��
�������� ������� ��������������� ��������
�� ��������� � ���������� ���������� ������
��. ��� �.�.�������� ��������, ��� �...���� �����-��
���������������� �������� � ��������� �������
������, ����� �������� ��������� �������������
��� ����������... ���������� (��������, 1974. �.186).
��� �� �� ���������� ��������� �� 100 �� 500��2
(��� ���������� �� ����������������� ��������) �
��������� �������� �������� � 1000��2
(��������, 1970, 1974).
�.�.�������� ������ ���� ����������� �����
������������ (��). ������������ ��������
������������ ������� �� ���� �����������
�.�.�������. � ���������� � �������� �����������
�������� ������� ������ �� ����������
(����������� ����; ��), �� ������ ��������
����������� ������ ������ �� (�.�.
����������� ������ � ��������� ������������
��������) � �������� ����� ������ (�����,1975,1982,
1987�,1994). ��� ����� �� ��� �� ��������������� �
������ ����������� ������������������
����������� �������, ����� ��� �� �� ��������� �
� ������ ������������� ���������������
������.(�������, ��� ����� ������ ����������
������ ������ � � ����������� ���� �����������.)
������ ������ � ������� � �������� �� ���� ��
����� �� ���������. ���������� ���� �����, ���
������� ������� �� ����� �����������
����������� ��� ������ ����������� ���, ������,
���������� � ������ �������� �����������
������� � ������ ������������� ��������. ���, �
�������� �-��� ���� ������� ��������� �� �����
��������� � 1000��2 (���������, 1987). �� ������
�.�.���������� (1987) ��� ���������� �����������
(������ ����������), ������� ��������������
������������ (������������� ��) ����������
200-300��2. ��� �������� ������������� �
������ ��� ���������� ������, ���, ������,
�������� ������������ �������������� ������ ���
����� �����������, ������ 100 ��2, ��� �
����������� 300 ��2 (� �������� 600-650 ��2
; ������, 1987).
������������ ����� ��������� �� ������� ��
����������� ������� �� ���� �������� �
����������� ���������� ����������� �������
������ �� ��������� ������� ������������
������� ������ ������ (������-������� ��). �����
���� ������ �������� ���� � ����� ����������
�������� ��� ������ ������, ��� � ������� ���
��������� �� � �� ��� ���, ���� ��������� �����
����� �������� (� ����������� ��) ����
�������������� ����� ����������� ����� �� ���
��������� ����� �������� �� ������ ��
����������� ������� �� (�����, 1987�).
�� ���� ����������� ��������� ��������������
���� ���������� ������� ������������ ��������
������ (� ������ ��������� ����������
������������ ��� ������ �������). ���������� ���
���� ��������� ����� (��) ����� ��������������
������������� ���� � �� ������ ����������
�����������. ������� ��������� ����� ���������
������ (��������, ������� ������) ����������� ��
������� ���� �������� ��. ������ ��� �������
���������� ���� ����� ��������� ������ ��
������� ����� �������� ����������, ������
������������� ������ � � ������� ������������.
������������� �� �� ����� ����������� �������
����� �� ������ ����� � ����� ���� ����������
������ ������. ��������, ������ ��������� �����
����������� ������������, �� �� �������
��������, �.�. ���������� � ���������� ����� ��, �
� ��������, � �������� ������ ����� �������
������������ ������� �������� ������ ����� ����
������ ������� ���������������� ���������
(�����, 1987�). ��� ���� ���� ��������� �������,
����������� ��� �������������, ��� � �� �������
��������, ����� ����� ������� �������, ������
��������������� ��������-����� �Ի. ����������
���������� �������� � ���� ������ � �����������
�������� ����������� ����. ������� � ������
�������� ������ ���������� ������� ������������
������� �������� ������ (�����, 1987�).
���������� ������ ��������� ���������
�������� � ����� ����� ��������������� ����,
���� �������� � ��������� ��������������
�������� � ����� ��������� ���� �� �����
�������������� ������� (�����, 1975, 1987�; �����,
�������, 1987�, 1987�, 1991). ����� �������� ����� ������
��� ������� �������������� ����� ����������
������ � �������� �� �� ������ ����. �������,
����� ������� ������� �� ����� ���������
�������� ����� ������������������� ����� �
����������� �� ������������ � �������������
�������� ������� (���������, 1987), �� ���� ����
������ ���������. � ����� �� ����� ����������
���� ��������� ������ �� ������� ������
� �������� ��. �� ��������� ������������ ���� �
��������� �������, ������ ��� ����������
��������, � � ����� ���� ������ � �������� ��
��-�������� �� ��������.
����� ������������� ������� ��� ��������� �����
���������� �����
�������� � ������ ������� � ���������
������ ��. �.�.����������� (1981�, 1981�) �
�.�.��������� (�������� � ��., 1994) ���� ����������
������������� �� � ��������
���������-�������������� ������� (���).
��������� ��� ���� ���������� ��� ������
������������� ������ � ������ ������������
����� �������������� � �� �����������,
�������������� �� ���������� � ����������
�������������. ��� ��������� ������, ��������
������� ������� ����� �������������� �
�����������, � ����� ����������� �
�������������, ��������� ������������
������������������ ������� �������� ����
������������� ����. �������������� ������� ��
��� �������� � �������������� �������������
(����� � ������������) � ������� ����������
����������� ������������� ��������� (� ������
��������� �����������) ������������ ���������
(��.�����: �������, 1995�).
� ����� ����������� �.�.����������� (1981�, 1981�)
����������� ��������� ������������, ��� �������
�������� �������� ��������� ��� �������
��������. ��� �������, ������, ��� ����
������������ �� �������� ��������� ����������
�� ��������� ������������� ������� ��� �������.
���������, �� ����, ������������ ����� ���������
������������ �������� ��������������, ���
��������, � �������� ��������� �������������
����. ������ � ����� �������� ��������� ��� �����
� ��� ��� ���� (����� � ������������) �������
���������� �� ����. ��� ���� ����� ����������
��������� �������������� ����� �������������,
���� � �������� (�������, 1995�). � ��������
����������� ���� ������������� ������� ���
������ ��������� ����� ��������� ���������� �
���� ��������������� (��.: �������, 1987; �����, 1992�)
� ��� �����, ��� ����������� ���������
������������ ����������� � ����� (Tansley, 1935; Dansereau,
1957, � ��.).
������� ������������� ������� ���������
������������ ����������� ������ ����������,
�������� ������� ������ �� ���, � ����� ��������
��������� ������������ ������ �����
������������ � �������� ����� ������ �������
(�����������, 1981�:154). �������� ����� ����������
��������� ��� ����� ��������� ���� ����������
��������� �������� ���������, ���������� �
������������ �������� ���������� ���� ��� �����
������������� ���� (�����������, 1981�). ��
���������� ����, ��� ����� ������������� �������
(� ���) ��������� �������� �����������, ������������� � ������ ��
�������� ����������� �.�.��������� (�,
��������������, � ������ �������������
��������������� ������ �� �.�.������). ������
����������� ���, ���������� �.�.�����������, ���
������� � ��� ��� � ������� �� ��������� ������
��������. ������ ���� ������� ���������.
��-������, ��� ����������� ������������� ����
����� ����� ���������� ���������� ��������
�.�.����������� (1981�, 1997) �� ��� ����������, �
������ ������������� � ������������. ���
����������� ���������� ����, �����������
����������� ���������� �������, ���
����������� � ������������� ��������������
����, ������������� ��������� ������� ����
������� ������������� ����������.
��������������, ��� �������� ���� �����������
����� ����� � � ���� ����� �������� ���������
���� ������� ������������� ��������� (���� ���
����� ��������; �����������.,1994); ����������
����� ��� ��� ������������ ����������
����������� �� ������������� ����, � ������ � �
�� ������� �� � ��������� �������������, ��� ����
�� ���������.
��-������, ������������ ��� ����� �.�.���������
(1962, 1974) ������� ���������� ������ �����
��������������� �������� �������� ������������
�� ������� �������� ������ ������� �������
(�������, ������, 1936; �����, 1956), �� ����� �� ��
������ �������� ���������� ���������
(����������� 1969, 1981�, 1999), ��� � ������������ �����
�������������� ����� �����. ���������� ���,
���������� ����� ��������, ���������� ������� �
(��� ������) ������ ����������� ���
��������-������������ (� ��� ������������
�������� ��������� �� �������������
������������ ������ ���), ��� � �������������.
���������� ������� �� ��� ���������� �������
����� ������ ������� ��� ���������, ��� �
��������� (�������� ������� ��������� � ����
��������� ���������� ����������). � �� �� �����,
�� ������ ����������� �.�.���������, ����������
�� ������ �� ����� ���������� �������
��������� ��������������� �����.
���� ��, ������, ���� �� �������� �������
�������� ������������ ������ ��������� ������
�� �� ������������ ��������� ������� �
���������. ����������, ������ �� ��������
������� ����� ��������, ���������������
��������� ��� ������������� �������, ��� � ��.
������ ����������� ���� �������� ��� ������� �
���������� ���� ������.
� ���������� ���������� ����� ���
������������ ��������� � ������������ ������
����
������ ������������� ����������
��� ���������� ���������� ����� �� �������
�� ���������� ���� ���������, ��� ���� ��� ��
�������� (��)����������������� ��� ������ �� �,
�������������, �� ���������� �� �������� ��
���������� � ����� ���������� �������. �����
����, ������� ��������� � ���, ��� �����
���������� ��������� �������� ����� ���������
�� (��� ������� ���������������) �� �����������
������������� ���������. ���� ��� �� ���, �� ��
����� ����� ��������� ���� �� �������.
���� ���������� ���� �� ������ � ����������
������������� �������, �� � �� ��������������
��������������� ������ ���� ����������� ��
��������������� ���� ����������, � ������, ��
������� � ��������� ���. ������ �� �����
��������� ��������: ������� ������� ������
������������ ����� � ������� ��� �� ������������
�������� ���������.
���, ����� ������� ������ ����������� ���� Agrostis
canina2 ��������� � ����� ��������
��������������� ��� Picea abies (�������� � ��., 1992)
�, ��������������, ����������� ���. � ��������
������������� ����� �������������� �����������
� ���������� ��� (�����������, 1999) � ����� ������
���������� Poa malacantha � Luzula beringensis,
����������� ������������ (���������� �
������������ ���������; �����, 1984�, 1991�; �����,
�������, 1993) ����, �������� � �������� � ��������
����������� ��������� ������, � ����� ��������
������������� � ����� ���������� ���������. ����
��� ���������� ����� �� �������-������������
��������� ����� ��������� ������������
�.�.����������� (1981�. �.203-204) ��������� ����������
�������, ��� ��� ���� ��������� ��������
�����������. ������� ����� �������� � � Potentilla
vulcanicola (�������, 1993), ��������� ���������
������, ������-�������� ������ ���������������
�������� �� ������� (������ � ���������
��������������� ������ ������ �����) ���������
������� ����������� ��������������� ������,
����������� ����� ��������������� �
������������ �������� (�����, 1974�), �. �.,
����������, ����� ���������� � ��������� ���.
�.�.����������� (1997)3 �������� �����������
������ ��� ����� ���� Rosa � �������� �������
�������.
����� �������, ��������� �������� �����������
������������� ��� �������������. ������, ����
���� ������������ ��������� ������������
��������� �����������, ���� �� ������� �������
��������� �������������� ���� �� ���������
(������� ������������� � ������������), ��� �
������� ���������, � �������� ������ �� ������
������ ��� ������.
������ �������� ���������� �������� �����
�������� ��������� ���� ��. ���� ���� ��
�������� ������ �� �� ����������� �����,
������� ���� ����������� ������� �����, �� � �
������� ��������� ���� ��������� ����� ����
�������������.
���������� ��������������� �������� ��� ��
���������� �������� ������������ � �������
���������� (��������� ������� ��������� �
�������� ������� � �������������), �������������
�� ���������� ����������� � ������������ ���
(��� ����� ��������� �������� �������� �����),
������� � ����.1. ���������� ���� ��������� (52%) ��
����� ����-���, ��������, ����������� ��������
�������� ��������� ��������. ����� ������ ��
��������� � ������� �� ���� ��������� �� �����
����������-������� ����������� �����������
������� ������������� �������
������������������, � ���������, �����������
������ ����������� ������ � ����������-�������
������ ����� ��������� ������� ������������
����� �� ��������� ��������� (������, ��������,
1976). ��� ���� ������ �� ���� ���������� ����� �
�������� ����������� ������� ����� ���������
����� 2/3 ��� ������������ � ����� 3/4 ���
����������� ���4.
���� ���������� ������� � ������� ��
���������� � ������ �� ��, ��� ����
�������������� �� ��������� �� �������
��������� ����� (�������, ��������, ������ ��
������ ������ � �� �������� � �� ������ �����
��������-���������� �����������, �������
������� ����������� � 7,3 ����). �������������
�����, ��� �������������� ��������� ���������
���������� ������� �� �������� �����
�������������� ����� ������������ �����������
������������� �������.
������� ������������ �� ���� ����������
������, ����������� ��� ���������� ��������. ��
���������� ������������ ����������� �
����������� ��� Festuca rubra, Juncus atrofuscus, Rumex crispus,
R.thyrsiflorus � Coronaria flos-cuculi, ��������
�������������� ������� ���� ��, ������ ����
����������� �����, ������������� ���� � �����,
�������� � �� ���� ���������� ��������������. �
�� �� ����� � ������� �� ����-��� � �-�� �������
��� �� ���� ������������ � ������ � ������
���������� ����������� �������� �����������
����������. ��� �� ��������� �� ����������
������ ��� � ������������ (�����������, 1981�).
����� ���������� ������������ ��������, ����
�������� � ����� ������� ������ ������������,
��� � ������ ������ �� ��������� ���������
�������� ���������. ����������, �������� ������
������������ Quercus robur � Corylus avellana �
���������� ����������� (���������� ���), ���� ���
��� ���� �������� ����� ������ � ��������. ����,
��� �������� ����������� ������� �� ���� �����
��� �������� � ������� ������� ���������������
�������������.
������������ ���� ���������� ����� �� �����
���������� � ��� ������������ ��
��������������� ���������. ������ ����.1
���������� � ����.2 � �������
������������-������� �������� ��. �� ����.2
���������� ����, ��� ��������� ���
��������������� ��������� ���� � ������ �������
������������ �� ���������� �����������,
����� ��� ��������������� ������ ���
����������� ������� ����������� ���������� ��
������� ��� ������ ������ ��. ���, ���� Asteraceae
� Scrophulariaceae ����������� ���������, � �� ������
����� � ������ ������� ����� ���������, ��� Ericaceae
s.l. (incl.Vacciniaceae), Ranunculaceae, Pyrolaceae, Equisetaceae,
Polypodiaceae s.l., � ����������� ����� Lamiaceae. � ��
�� ����� ������� ����� ��������, ��� Poaceae, Cyperaceae, Caryophyllaceae � Rosaceae, �
����� ������ ��� ����� ������� �����.
������������ ���� Brassicaceae � ����� ��������
����������� �� ������������ ����� ����������
������������ �����. � �� �� ����� Orchidaceae � Salicaceae
�������� ������ � �������� ��� ��� �����������
������� ��, ������ � �����, ��� � ��� ��������� �
�����������, ������ �� ������ ������� �� ����� ��
���.
����� �������, ��������� ���������������
�������� ���� (�������������, ��� �� �����, �
��������-������������, � �����������) ��������
�� ���� � ������� ������� ����������� ���������,
� �� ����� ��� ��������������� ������� ���������
�������� ����� ���� ��������������, ���������
������������ �������.
������ �������������
���� �� ��������� ���������� ����������
����� ��� ���������� ���� �������������
���������� (��� ���� �� ����� ���� ���������), ��
������ ����� ��������� � ��� ���� ���
����������� ������������� ���� �, ���� ��������
� ������ ����� ��������� �.�.������������,
��������� ������� ������������ ������������ ��
����� �������������.
����� �.�.����������� (1981�, 1997) ����� �
����������������� � ��������������� ����
���������, � ����� �������� � ������ ���������.
��������������, ��� �������� �����
�������������� ���� ����������� ���������
(������������, ������������� ��������; �������,
1993). ����������, ������, ��� ���� ����� ��������
���������� ������� ���� ����, ��������������
��������������� ����� ������ ���� ������������.
��������, ��� �� �������� ��� ��� ����������
����� ����� �������� ������������� ���������,
��� ���� ��� ���� �� ����������. ���, �
��������-��������� ����������� ������������
������ ����� Alnus glutinosa �������� ������
������ ����� A.incana, ��� �������� ���
�.�.�������� (1975). ��� ���� ��������� ���� ������
�� ������� ������ ������� (�������� �� ���� �����
������ ������ �����), ������� ������� �
����������������.
���� ������������ �������� ����������
������������ ��������, ���������, ��� ���������
������� ������������ ��������� � ����
������������� ����� ������������ �����, ��� Aegopodium
podagraria � ���������������� ����� �������
������� (��������, 1987), ��� Calamagrostis epigeios � ����
�� �������� ��������� �������� �������
(�������, 1980) � � �������� �����. ������ � ����
������ ������ ���� ������������� �����������
�����, �.�. ��������������������� ������
��������� �����������.
������ ���������� ���� (��� � ������ 90% �����
��������) ����������. ����������� ���������, ���
����������� ������-��������������������� ��
�������� � ��������� �������������; ������� ���
����������� �� ����� (����� �� �������� ����
��������, ������������ � ��������� �����) ���
����� �� ����������� ����������� ��
���������������� ������������. � �� �� ����� ���
��������� ���������� �� ����� �� �������
����������� ����� ������������� �������������
��� ��������������� (Harley, Smith, 1984; Bagyaraj, 1991).
�������� ������ ���������� ������� �������� ���
��������� ����������� ��������� ����������, �
����� ��������������� �����������
��������������� (Harley, Smith, 1984), ����� ������������,
��� ��������������� ���������� ����� �������
��������� ��������� �������� ����������
���������-���������� ����������� ����������
(���� ��� � �� ������� ������� � ���������
��������� ������ ���������� �� ��������� ������
�����). ��� ������������� ���� �������������
���������� ��������� ������������ �
�������������� ��������� ������, �� � ������
����, ��� ��������, ���������� �������������,
����� ����������� �������������� ��������� ����
��������� �, �������������, ������ ��
������������ ��������� �����. �������, ������,
��� ��������� � ������ �������������
��������������� ��������������� ����� (����
��������� Alismatidae, �������� Juncaceae, Polygonaceae,
Chenopodiaceae, Caryophyllaceae, Brassicaceae, Saxifragaceae; ���������,
1981)5.
�������, ��������� ��������� �������
��������������� �������� ������ (��������� �� �
���������� ���������; �����, 1984�, 1991�; �����,
�������, 1993), ��� ��������������� �
��������������� ��������������� ��������������
�� ��������� (��������, ����� �������������
��������������). ���, �� ������-������ ������
���� ����������� ����������� Carex echinata,
����������� ��� ������ ���������� �������� (Deschampsia
cespitosa) �����: ��� ����� ������������� ���� �
����� �������� ������� (�.�.�����, ����.�����.)6.
����� �� ��������� ������������ (������
���������� ���� � ������� 4�� �������) � �����
������������ Poa remota � ������������
�������������� ���������� ��� (����������
������ � 1993�.).
��� ��������� ���� ��������������� � ���, ���:
- 1)������ (���� �� ���) ���������� ���� ������
������������ (����� ������) ���� � ���������
����������-�������������� � ���������������
����������� ������������� �������, � ������ � ��;
- 2) ���������� ������������� ������, ����
��������� �������� ��� ����� ���������� ����� �
��� ����, ������� ��� ������, �����������
�������� ������������� ��� �
�������-������������, ��� � �� ��������������
���������;
- 3)���� ���������� ���� ���� ����������� � �����
������ ������ ������������� ���������.
�����������, ������ �����, �� ��������� ������.
����� ������� ������������ ����� ��������
����������� ������ ������������ ���������
���������� (� ������� ������) ����� �� ���������
����� ��, �� ��������, ��� ��� ����� ����
���������. � ������ ���� ������ ��������
�������� ����� ��� ����� ������������ �����,
������ �����������, ���� ������� � ���������
��������������� ����������. ���� ��� ����
����������� ���������������; � ����������
����������� � ������������ ����. � ������������
� ��������� ����������, � ������ ������������
��������� ����� ����� ����� �������
�.�.������������. � ����.3 ��� ���� ����������
����� ���� ������� �� ��������� �����
������������ � ������ ����������� (�� ������
������� ������������ ������ � 1982-1993 ��.).
�������������� ��� ������� �� ������ ������� ��
�������, ������ ������������ ������ ���� �����
���� ��������� � ������ ������ ������ (��.����).
��������������
����������� ���� ������������� ��������,
������� � ������� ������� ������������� � ���. ��
������������� � ����������� ��������������
�������� ���������, ���������� ������������� ��
����� ��������������� ������������ ���������,
������ ��� ���� ������� �� ������ ��� ����������
��������� ������� � ����������� ����� �����.
�������� �������� ��� �������:
��������������-���������
����������� ��������� ����, � �������������
�������� ��������� � �������������. �����������
�� �������� ����� ����������� ��, ��� ������
����������� ��� ������� ��������� ������������
(������ ������������) ���������. CR-���� (Grime, 1979)
������ ���� (Typha latifolia), ��������� ����� (Petasites
spurius), � ���������� ������ � ��������� ������
�������������� (Potamogeton gramineus, P.perfoliatus, Myriophyllum
spicatum s.l., etc.).
��������������-�����������
������-����������� (S- � SR-) ���� (Grime, 1979), �����
����������� ��������� � �������������; ������ �
���������������� �.�.������ (1986, 1995).
������������ ������ ������������ (���
������������ � ���� �������, ����������� ���
����� ���������� ���������), ������ ���� ��
���������� ������� ������ �������������:
��������������� �� ���������� �������������
�������. � ����� �������������� � ������
���������� � ���������� ����, ������ ���
����������������� �������������, �������� �
�������� ������ ����� (�����, 1968, 1974�). ������
������ �������� ���������� ��� ������
���������� �������, ��� � ���� ������� �������
����������� ���� ���� ������� ��������� (Phippsia
algida), ��������� ������ (�� ������� Claytonia arctica,
Saxifraga eschscholtzii, Senecio jacuticus, Crepis nana) � ���� (� ���
����� ������ ��������� �������� ���������). �
������ ������ ����������� ����� �������� ������
�������� (Salicornia pojarkovae �� �������� ������
����), �����������-�������������
����������-������ ����. ����������
������������� ��������������� ��������, ����
���� � ���������-����������� ����, � ��� ����� ��
�������������� ������ �������� (Lobelia dortmanna;
Harley, Smith, 1984).
����������
����������� �.�.���������� (1938), ��������
��.������ (Grime, 1979). ��������� �������������
����������� ����������� ���� �� �.�.������������
(1981�). � ����� ������ � �������� r-�������� �
���������� � ������������, ���� � ��������, ������� ��������������
�������������, ���������� ������� �������������
���������, ������ ������������ �����������
������ � ����� ������������ ������������ �����.
���� ������������� � �����������, �� �� ������� �
����� ��������, ������� ������ � ����� ���������.
���������� ������ � ����������� ������� �
������� ������� ������������� � ����������
�������������������� ����������. �����
���������� ��� �������:
����������-���������
�����������-��������� ����, ������� ����������
������������ ��������� � ������� ��������
��������������� (Urtica dioica, Chamerion angustifolium, Cirsium arvense)
� ������ ����������� ����������� �������
�������� ���������. R-, CR- � ���� �-���� (Grime, 1979).
����������-�����������
����������� � �����������-�������������
(�������-, �������������) ������������, ���������
������� �������� ��������� ����������� �������
��������� ��������� (Capsella bursa-pastoris, Taraxacum officinale)
��� ����������� ������� � �������
�����-���������� (Rumex crispus, Cerastium holosteoides, Anthriscus
sylvestris, Androsace filiformis). R- , CR-, SR- ���� (Grime, 1979).
����� ����������� ������ � ��������������
������ (������ Brassicaceae) ������� ���� �
�������������� (Poa annua) � ���� ���������� (Plantago
spp.; ���������, 1981) �������������.
�������� �� ����� ������������� �������������
����, ����� ���������� ��������� ��� ���������
��� ��� ���� ����� � ����������� ����
����������� �������. ���, ��������� ���� �������
���� ������������ ������ �� ������� � ������
������� ���, ��� ���� ������ ����������� ��
������������ (Echinochloa crusgalli, Poa annua, Fallopia convolvulus,
Erysimum cheiranthoides, Descurainia sophia, Plantago major, Sonchus arvensis s.l., Cirsium
vulgare, etc.; �������, 1958; ��������� � ��., 1987;
�������� � ��., 1992). �������� ����� ���� �� �����
������������ ��������, �������, ������
������������ (��� ���� �� ��� �������
�����������) � �������� ������� ����� ������
������, �������� ������ �������������� (������
����� �������) � ��������� (������ ������ ��
�������!) ��� ������ (Chelidonium majus, Tanacetum vulgare � Crepis
tectorum � ����������-������ �����������; ������,
��������, 1976). � �������� ������� ���������
����������� ������ �������� ���������, ��������
������ ����������, � �������� ������� �������.
������ �� �����������/����������� �������
������� ���� ������� ������ ��� ������ �� �
�����������.
������������
����, ����������� ������� ���������
���������, �� ��������� ������������ � ���������
� ������� (��������������� �������) ������� ��
��������� ������� ���������. ��������������
����; �� ���������� (��� ��������� ���������� ��
������������� �����) ����� ��������� ��-�������
(����������� ���� �������������). ������ ����
�������� � ���������� ����������� (Dianthus fisheri,
Knautia arvensis, Campanula patula, Leucanthemum vulgare). � �������
��.������ (Grime, 1979) �������� � CSR-���������.
��������������� �������������-��������������.
�������������� (���������-����������)
�����������-���������, ����������� ������
����, ������������ �������� ������ ������,
������ ������ ���������. �������� �� ������
���������� ��� ������� �� �����, �� � �������
���������� � ������������ ������������ ������ (Calamagrostis
epigeios, Aegopodium podagraria). �������������;
���������������� �� ������������� ����� ���
������ �����, �� ���������� �� ����� ������ �����
���������� ����������� ������. �������������
�-���� (Grime, 1979). � �������� ��������������.
��-, ����- � �������������� ����� ��������� �
�������� ������������� ����� ��� �������������
(��.����), � ����� ����������� ����� ���
�������������� ������������� �� ������
�����-������ (��.: Sumina, 1994).
������������
����, �������� � ������ ������������
������������ ��������� ����������
������������� �����, � ������ �� ��� � �����
��� �� ����� ������. �������������� (����
���������������) ����; �� ����������� ����������
� ������ ������, �� ������ ������������� �
��������� �� ����������� ������ ����������.
��������� �� � ����������� ��� �� ������
���������, ��������� ��� ����������� ����
�����, �����, ����� ��� �����; ��������� �� ���
��������� � �������� ����������� ���
����������� �������� �������������� �����.
����� ����� ������� �������� ��� �������:
������������-�����������
������ ����������� �����������-��������� (Pteridium
aquilinum) ���� ��������� (Sorbus aucuparia) ����.
�������� �������� ����������� ���� (������
�����) � ������ �� ���������� ���� �������
������������ ������; ��� ���� �������� (� �����)
��������� ���������� � ���� � ������������
������������ ����������� �������, ���� Piceetum sorbosum
�.�.������� (1939) ��� ���������� ��������� Pteridium
aquilinum (Watt, 1947, 1955). C- � CS-���� (Grime, 1979). �������
���������� � ����� ���������������.
������������-�����������
��������� ��������������, ���������� �������
�������� ���������� ����������������� �����
�������������� (��������� ����������� ������,
��������, ����������� ������� ��������� � �.�.).
�.�.������� (1965) ������� ����� ����
�������������� ������ ���������� ������
(�������, ����� �������������� ���������� �
������ ���� ���������, �� ���� �������� ��
������� ���� � ������������� ��������, �������
������������; �������, 1996). ��� ����������
��������� ��������� ������ ���� �����
����������; � ���������� ��� ��������
�������������� ������ �������7 . � �������
��.������ ��� ����� �������� � �������� � CSR-����
(��.����.3). ��������, ��� ��������� �������������
�������� �������� � ���� �������������.
������������� ���� ����� ������������� �
������ ����������, ��� �������. ������ �
����������������, ���������� ����- (Salix spp.) �
������������������ (Moneses uniflora) ����, � �����
�������� �� �������������� ������ �������� (����
Orchidaceae).
������ ������������ ����� ����������
�.�.��������� (1963) �� ���� ��� �����������
��������� � ����� ������� ������; ����� (� ���� ��
���������) ��� ����� �.�����-������ � �.�������
(Braun-Blanquet, Pavillard, 1928). �.�.��������� (1987) ������
�������������� ������������ ��� ������� ���
������������� ������ ����� (�������������;
�����������, 1981�), ������ ����� ��������� �������
���� ��� �� ����� �������������������.
�����������
���������� ��������� ������������
��������������; ��������������� � �����������
����, ����������� ��� ���� ����������������
������ ����� ������������� �����. ���
�������������� ������������� ���������
�����������������. � ���������� ���������
�������� �� ��������, ��� ���� �� ��� �
����������� �����, �� ������������������ ������
�� ����������. ��� ������ ���������� ���� ����
������ ����������� ������; � ����������
����������������� �������� ��������������
���������� ���������-����������
�����������������.
� ����� ����� ���� �������� ������
��������������� (Braun-Blanquet, Pavillard, 1928). ������
��������� �� (��� � ���������) � �����������
������������ ������������ ���� ���������
��.������ ��������������. ����� ���� ������� �
������������� ��������� � ������� ����������
��������������, � �������� �������������� �����
�� ���� � ������������� �������� �� �����������
���� �������������� ������ ������.
�������������� ����� �������� �������� �
������� ���������� ��������� C-, S- � R-��������� �
���������� ��������� �������� ����� � �������
��������� ������������� ����� ���������,
������������ ������� (Grime, 1979. P.73-75). ����������
����� ��� ��������������� ��������:
���������� �����������
����, ����������� ��� �����������
���������������� ���� (�������� ������ �
���������).
1. ��������������
��������� ������� ��������� � �������������
���� � ������� ��������������� �����������.
�������� ����������� ������ � ������ ����� Alnus
glutinosa, ��������� �������� ����� ����� A.incana
�� ����� ������� ��������������� ���������8.
��� ���� ������������� A.glutinosa ����������� ��
������ �� ������ ������� ��������� ����������,
�� (�� ����� ����������� � ��������-���������
�����������) � ������ ���� �� ���� �� �������.
������� ���� � ����������� �� ���� ������� ����
(������, ��������������, � �� �������������) �
�������� ��� ��������� ������������� ����� Salix
�� ������� ���������� �-��� (�����������, 1989).
2. ��������������
������������� ���� ������������ �������. ���
����������� ������� ���� ��������� ���� ������ �
���� ������: ���������� � ��������� ����������
����������� (��. ����.3); ������� ���������
������������� ��������� ����������� �����
�.�.��������� (1975, 1986) ��� �������� ������������
���������� �����.
���� ������ ������ ��������������� ����������
� �������������� � �������� ��������������: ���
�������������� ���������� ����� ��������
������������� ��� � ������� ��������� ���������,
��� � �� ������������� ����������� ������ (Veronica
chamaedrys, Galium album). ��������� �������� ������� ���
����� ����������, � ����������� ��� ��� �������
������������� �������������� �� ��������
������. ������� ������ ������, � ������
������������� ���� ������� ������� �����������
����, ����� ��� Jasione montana ��� Artemisia campestris.
����������� ����������� (������
��������������)
������������� ����������� ����, �����������
��� ������ ���������������� ������ ������
�������������� ����: � �������, Linnaea borealis � Gentiana
pneumonanthe �� ��������� ������ ��������������
����������� � ��������� ���������� �����������
(��.����.3). ��� ����������� ��������� ���������
����� (��������, Populus tremula) � ���� ������������
����� ���� (Carex pilosa), �.�.����������� (1981�)
������� �� � ���������, ������� � ������� ������
�� ��������������� ������ �������������� ����.
����������� ������ �����������
������������� ����������� ����������� ����
����������, �������� ��������� �.�.�����������
�������������� � ���������� �� � ��������
���������� � ����� ���������.
��������
���������� ������� ����� �������������
������� (��� ������� � ������ ����), ��������
������� ��������� ���� �� ����������. ��
�������� ���� ��� ���, ��� �.�.�����������,
������� � ���� ������ ����������� ���������
������������� ������. ��� ������� �� �������
�������� ������ � ������������ ��� ����������
����������� �������� �� ������������ � ������������
(� ������ ������������ ����� ������������ � ���������������
� ���������������) �������� ��� ����
���������, � ������:
���������������
��������� ����������� ��������� ������
��������� � ���������, �� ���������� � ������
������ ������������� (Festuca ovina, etc.). ��� ���� �
����������� ������, �� � ������� ������� ����
���� ��������� ��� ������� ���������
������������ �������, �������� ���������
������� ������������ ����������. � ���� ������,
��������, ������� ������� � ��� ����������
��������� �������������� (��������, Cassiope tetragona
��� �������� �������������� ����� ���������
����� ����� �������); �������������� ��������
��������� �������� ������ ������������ ������ �
��������� ��� ��������� ����������� ��������9.
�� ���� ����������� ��������� � �����������
������� �������� ���������� ���� � ��� �����,
������� ������ ��������� �������������� (���
��������� ��������� ��������������� ������).
���, �������� �����������-�������������
�������� ���� �������� � ����� Carex obtusata, ��
����������� ������� ������������ � ��������
������������ ����������� ���������� � ���� ����
��������� ���������� ������ (�������, 1996).
���������� �������� � (���)����������� ����� �
������ ������� ������, ��������� ���������
������������� �������������.
����, ����� ������������� ���� ����� ������
���������� ������ ������������� ���������
�.�.������������ (1981�, 1997), �.�. ����������� ������
����� ���������� ��������. �������������� �����
�� ���� ������ ������������� � ������
������������� �������, ��������� �������������
�� ������� ����� ���������. ������������ ���
�������� ���������� ���� ������� � ������������
������������ �������, � �������������� �����
���� ���� ����������� ������ ���������.
������������ � ����������� ��������� �������
���������� � ���������� ��������������, �� ��
����� � ���� ���������� ����� � �����
������������ � ������� ������������
��������������. ��� ����������� ����� �������
������������� � ������������� �������
���������� ����� ������������ �� �����������, �
��������������� ������� ��, ������, ������������
(� ���������� ���������, ����������� ���
�������� �� �������, ����� ���������� � ����).
����������� ������������� ��������� �������
������������� � �������� ����������
������������� ����� ��� ������. ��������� ������
�.�.����������� ������� ��� � ���������.
��� ����, ����� ������������ ��� ���� �� ��
����� ������������ �����, �� ���������
���������� ���������� ������� ���
������������� ���� ��� ����������. ���� �����
�� �������, �� ��������� ���� ��������
���������� �����, �������� �������� ������
������������� ����� ���� � ���� ����� �� ����� ��������������
������� (�������, 1995�). ���, ��� �������
��������� ����� � ���� ���� �����������
�������������� ����������� ������� ���������
��������� ������: 1) ����������� ��������������
���� � ����, � ������� �������� �������������
�������������� �� ���� ��� ������� �
������������ ��� � ������ �����: 1�)
�������������, 1�) (�,�) � ���������� � ���������
(�������������, ���������) ������ �����, 1�) �
���������� � ������������� ���� ���������; 2)
��������������� ������ ���������, �����������
�� ����� ������ ����� � ���������� �� ������ (���
2� �� ����������� �����������): 2�)�������, 2�) ����
������������� ������, 2�) ���������; 3)
����������� (� ������� ������) ����, ��
����������� ������� �������������� � ����������
������ ���� (��������� �� ��, ��� � �
������������); 4) ��������� ����. ���� ������
(���)������ ����� �������������� �� �������
�������������. �������, ������, �������, ���
������ ���� � ������ ������������� ����� (���� �
�������� ������ ���������) ����� ���� ���������
(�������, 1996).
��������� ���� �������� ������������ �����
���� �� ����������� �������� � ������������
����� ����� (������� ����� ���� �������� � ������
��������, �.�. �� ������ �����������, � ������
�����, � ��������� �������������� ���������), ��
�� �� �� ������������ ����������, ���������
��������� ����� ���� � ��� �������������� �����
��������������� � (�������) �������������
������������� (Grime, 1979). ������ �������, ��� �����
�������� ���� ����� ���� ���������� ���
��������� �����, � �������� ����� ������ ����� �
������ ���� ������� � �����������
�������������� ��������������� ���������.
������ � ��������� ��������������� �����
���������� ������ ������������ ���������, �����
������������, ��� ����������� �������� ��������
�� ������� ��������� ��������-���������� (���
������� ���������� ��������� �������
������������� � ������������ �����), �������
��������� �������������, ���������� �������
���������� ������ ������ ��������� ������� ����
� ����������� ����� ������������� � (�������)
������� �������������� ����������� �
������������� � �������� �������� (�.�. ���
������� ���������� ���������� ��
��������������� ��������) ����������������
������. ��� ������ ��������� ��� �������� �
������� ������� ������������ ������������� (�.�.
���������������� � �������������) �
������������ � ��������� �������� ��������� (Van
Valen, 1977; �������, 1987), ��� � ����� � ������� �
�������� � ������� ����������������, � ���������
� ���������� � ��������� ����������-�������� �
���� ��������������.
���� ��������� ��� � ������ ���� �����, ��� �
������ ���. � ����� ��������� ������� �������
������� ������������ ����������� ���������
����, ����� ��� ������������ � � ��������
����������� ��������. (�������, � ��������� �����
���� ������� ������������ �������
���������������). � �� �� ����� ��� ������� ���
����, ��� � ������� � ����������� ���� �� ��
����� ������� ����������� �� ����� � ���� �
�������� �������������� ��� ���������, ��� �
����������� ������������ � ������ �����
����������� ���������, ����� ��� ���������� �
������������ ���������� ��������� ����������
(�������, 1995�). �� �� ������� ��������, ��� ������
�� ������������ ����������� ���� �����������
��������������� ������������ ��������� � � ����
������ �������� �������� � ������ ��������
������������� ��� ������� �������� �������
�������. ���������� �������� ��������� �
�������� � ������������� �� ��������� ������
���������������� � ������� ������ �����
�������� (���� Poa, Ranunculus � �������� Carex).
�������� ����������� � �������� �����
����������� ������������� ������ ������������
�� ���.1. ������, ��� �������� ����� �������� ���
������������ ��������������� ����� (���������),
��� � �������������� ������������ ���������
���������� �����: ������������, ��� ������������
����� � �����-�� ���� ������������ �������
���������������.
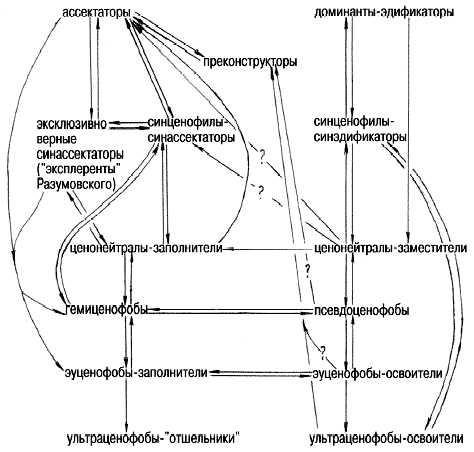
���.1. �������������� ����� ���������
������������ � �������������� ������ �����
������ ������������� ��������� ����������
��������
��������� ���������� �����
����� ��������� (� ������� ������) ���������
� ����������� ��������� � ������ ���� ����� �
������������ �������� � ����������������. �����
��� �� ������ ��������� (� ��� ���������������
������������� ��������� � ������������� �����),
������ ����������� ���� ����� ������� �����
������, ������ ��� � ���������� ���������.
���������� ����� �� ����������������� ������
������������� ��������� ����� ������������. ��
��������� ������ ����� ������ ����������
����������� ������������ ��������� ����������
�������������� (�������,1994), ���� � ����������
������� ����� � ����������� �����
��������������.
��� �������������� ������, ����� ����������
����� � �� ������������ �������������
������������� ����������� ��������� ���
�������� ������������ ��������� � ��������
����� ���� � ������ (�������, 1954; ������������,
1964), ������������� �� ����� ����� �����
������������� ����� (��������, 1964), ����� ��� � �
�������� (�������, 1954).
� ��������������� ����������, ������, ���
������������� ��������� ������������ ����� ���
����������� ������������ �������� (��������, 1964):
��������� ������� ������������ �� ����
���������� �� ���������; ����������� �����
������ ������� � ������������ ������
���������; ���������� �������� �� ��������� ��
�������������� ���������� (������) �
����������� ������ ����� ���������. ���������
����� ����� �� ������������� � ��������� �������
����� ����� ������������� ��� ���������, ��� �
��������� ���� ���������.
��������, �������� �������� ���������
������������ ��������� ������������� ��
������� ���������� ����� (����� ��.: �������, 1994);
����� ��� ��������� ��������, �������������
���������� ��������� ������������ ����������
(���� � ����������� ������� ������������ � ����
���������� � ����������). ����� ��������
(���������� ��� ������� ��������� ���� �����
����� ��� ����������) �� ���� �����������
��������� ������������� ��������� � ������
��������������� �� ��� ��������� ���
������������� ����, � ��� ���� �� ����������
�� ����������, �� ��� ����������� ��������
�.�.��������� (1983).
� ����� ������, ��� �������� ���������
���������� ��������� ��� ���������������
(�������������), ��� � ���������������
(������������) �� ������������. �������� ������
�� ��� ������� ��������� �������������
����������� ������������ ���������������
�������. ������ ������������� ����� �� �����
������������� ��������� ����� �� �����
���������� � �������, �������� �� ����, ���
����� � ����� ������ ������������, ��� ���������
�.�.����������� (1981�). ��� ������ � ������
�����������; ��� �� ������� ����������
�������������� ���������� ������� �������
������������� ��������������, � ����� �
������������� ����.
��������, ������, � ��������� ����������. �
������ ��������� ��� (�����������, 1981�, 1981�), ������ ������ (� �� � ����)
������� ����������� �����, ��������������� ���
��� ���� ������������ �����, ����� ����������� �
������ ������ ���10 (�� � ���� ������� �����
����������� � ����������� ������������ �� ��
������ � �� ����� ���������). ��� ��� ���
���������� ������������ ���� ����� (�
��������������� �� ������� ����������� �����)
������� ��� ���� ���� (�, �������������, ���
������������ ��������� ��), �� �� �����
������������� ������, ������������� �� �������
�����-�� �� ���� �������, � ������ ����� � ��� ��
��. ���� �� ����� ����������� �������
������������� ������ � ��������� ��, �� ������
������� � ������� ������������� ������ �������
������� �������, ���������� ������� ��������� �
����� ������ ��������� ���� ����� �����.
� ���������� ������ � �������� ������� ��
(���������� ��� ��) ����� ������������� ���� ��
����������, ������� �� ���������� �������
��������� ��������������� ���� �� ���� ������
�� ����������� �����. ����� ���������������
����� ����� ������������ ����� ������������
���������� �� ��� �� ������.
��������, ��� ����������, ���������������
����� ���������, ������������ �������� ��
�������.
�������������� ������� � ������������ �������
���������� �������������� �������������
����, ��� ���� � �������� ������������ ������
������ ���� �������� �������� ������
������������� ��������� � ������ ��� ������. ���,
Festuca rubra � Succisa pratensis, �����������
��������� ������� ���������� �� ����������� ��
����, ���������� ������������������� �����
����� ��������� ��� �������� �� ������� ��� �
������ ������ (�����������, 1981�), � ������������
�� �������� ��� �� �������� ��� Sorbus aucuparia s.l. �
�������������� ������� ���������� ���������� �
��������� ����������� �����������.
������������ ������ ��������� Anemone nemorosa,
������������ ������������� ��������� ��������
��������� ������� � �.�.������� (1930). ������������
���������������� � ���� �������������� �
���������� ����� �����������, ��� ��� ����������
�������� � ������� ���������������, � ����� �
�����-��������������� ����� �� ��������
���������� �� ������ � ��������� �������� ��
����������. ����� ��������� � �������-��������.
� ������ �� ������ A.nemorosa � ����� ��������
����������� ���; �� ����� �� ������ ��� �� �
���������������� ��� ���������, � �
����������������� ������ (��������, �������� �
������-���������). �� ��� ������ ���� �� ���
������� ��� �������� (������, 1983). � ������� A.nemorosa
���������� ����������� � ������ �������� �
������ �� ���-��������� ���������.
��������� ������������� ��������� �����
����������� � �� ����� ��������� ������������.
���, Carex cinerea, ����������� �� ��� ���������
�-��� � � �������� ������� ������ �� ����, �
������������� ����������� ������������� � �����
� ������ ������� ������������ �� ������������
��������� ��������, ��� ������� ����������
������������ ������� � �� ���������� ���������
����� ����� (���������� 1981 � 1993��.).
���� ���� ��� ������������� ��������� ����
�������� ����������, ��� ������������� ������
(������������ �����) ����� ���� ��������� �
������ ������ ������. ���� ��������� ����� Populus
tremula, ���������� ����������� ���������������
����� �� ������� ������ �� ������ � � �������
������ ����������� ������, � ��� �� �������
������ ��� �� �����, � ����� � ��������� ��������
������ �������� �������� ����������: �
�������� ������� � ����� ������������ ���������
���������� �������; � ��������� � �������
�������������� ��������� �����. � �������� ��
�������� ����� ��������� �-���, � ����� �� ������
�������� ������� (��������, 1985) ����� ������
������� ������ ��������, �������� � ������� �
������� �� ����� ������������ �����.
�������, ��� ������������ ��������� �����
����� �������� ��� ����������� ��� ��������
������������� ���� ������������������ �������,
��� �������� ����� ������� ���������� � �������
������ �����. ������ �������� ������������, ��� �
� �������� ������������� ���������� (��
�������� �������) ���������� ����� ������������
��������������� ��������, ��� �������� �����
������� ������ (��� ���� �� ���������) ���� �����
�������� ��� ������ ������������� ���������.
���������� ���������� ����� ����� ���������
��������� ���������� ��������������� �, ������,
�������� ������ ���� �� ����� (���� �� �����
������) ���������� (����� ������, ���������
������� �� ��� ���� �� ���������� ����� � ������
�� ����������� �������.)
��������� ������ � ����� (�����, 1982, 1987�, 1994)
������������ �� ������ ����������� ��������������
�����: �������� ������� ������ ���������������
� � �������� ������� ����� ��, ���� ����
�������������� ������ �� ��� ������� �����
������� ����� ��������� (��� ������������
��������, ���� � ������������). ����� �������,
���������� �� �� ������ ���������� ������� ��
������ ������� �����, �� � �� ����������� �
������ �.�.������� (1965). �������, ��� ��� �������,
��� ������������� �������� ������ ������
�����������.
������� ����� ��������, ��� ��������
��������������� ������� ����� ��������� ��
����������� �������, ���� �� �� �������,
������������� �����������. ��� ����� ��� �����
����������� � � ��������� ������� ���������
�����11. ����� �������� �������� �������
����� ������� ����� ����� ��������������
��������� ����������� (� ������� ������) � ������
(������ � ������������ ������ �������� �.���� ��
������ ��������� � �����������). ��������� ����
����� � ��� �� �����, �� ������� ��������� �
������������ ��������� ������ ���� �����, �
������:
- Matteuccia struthiopteris (����� ����������� ���� �
��������� � ����������� ����������� ���� ��
������ �������);
- Equisetum sylvaticum (��� ���������� �������
������������� � ��������� ���������� �
������������ ��������� (C(f); A3(b); He9,10,11(b-e),12(a-g)), ���
���������� ���� ����������, ��������� � ���� �
���������� ����������� ���������� (Hm3,4,5(b-e)), ���
���������� �������� �������� � ��������, ������
�� ����, ��� ��������-�������������� ��������
��������-��������� ��� ����);
- Lycopodium clavatum (������� �� ���������������� �
����� �������� � �������������; ���������� ���
���� ���������������� � �� ����������� �������
������������ ����);
- Milium effusum (������� �� ������������� �
������������ ��������� � �������������);
- Lerchenfeldia flexuosa (������� �� ������������ �
������������� (X5(a-f); C(a-c); Hm4,5(a-d));
- Carex pilosa (������� ��������� ���������������);
- Polygonatum odoratum (� ����������� ���������,
���������� ���������� ������� ��������
��������� �� ����� �������� �� ���������
���������� �����; �������� � �� �����������
���������� ������ � ���������� �����������
������ ����� ����� ��������);
- Alnus incana (���������� � ���� ������� ���
������� �������� ������������ ���� �� �������,
�������� �� ������������� ������; �������� ����
� ��������� ���������� �����������);
- Anemone nemorosa (��. ����);
- Trollius europaeus (������ ����������� ����������
��� ����������� ������������ ����);
- Prunella vulgaris (������� �� ���������������� �
��������� ���������� � ���������������� �
���������� � ������ � ��������� �����);
- Trientalis europaea (������� �� ���������������� �
������������� � �������������; ��� ���� ��������
���������� ��������� ������ �����������
���������� Picea abies � Maianthemum bifolium (Trientalis europaea)
����������� �� ��� ���������� ������: ������ Picea
� Trientalis, ����� Picea � Maianthemum; ����� � ������
��������� ������������ ��� �� � ����, � � ������,
�������������� ��� � ���� ���������);
- Linnaea borealis (������ ����������� ���������� �
����������� ����������� � �����������
����������). ��������� ���� � ������ �������, ��
�������� � ���� ������ ������.
����� �������� ������ �� ������ ������
�������������, ��� ��� ��� ������ ����������� ��
������� �� ���������� ��������������, ������� ��
������ ������������ ���������. ���������
��������� ������������ ���������������
��������� �� ��� ������� �������. ������, ���
������� ���� ������� ���������� � ��������
��������� ��������� �������������� � �
���������� ����������� �������� � ���������� ��
(������ ���� ���������������� ���� �Ի), ��������
������������ �� ������������� ����� ���� �����
������ � �� ������� ���������. ���
��������������� ��������� ���������
��������������� �������� �� �������� ���������
���������������� ������ ������ ��� ��������
����� �� ��������� �������, ��� �
��������������� ������ ����� (� ��� �����������
� ������������ ������������������ ������; �����
���������� ��� �����-�� ����, � ����� �����
��������!).
� ����� ���������� ����� �������� �������
�������������� �������� ����������� ����
��������������� ����� ������ � �� ����������
���������� � ������������ �������� ��������.
���������� ������ ������������� � ������
�������� ���� �������������� ���� ��� �����
����������� ��������������� (�����, 1992�).
��������� ������� ���������� ���������� ����
�� ������ �.�.������ (1932), �� �����
��������������� ����� �������������
���������������� ������ (���������� �
���������� �������� ��������� ������������
���������). � ���������� �� ������������ � ������
��������� (��������, �������, 1969) � ��������
������������ ��, ��������� ���������� � ��������
��������� (=��) ���������� � �����������
������������� ����� �������������� � ������
����������� ������ ����������.
��� ��� ����������, �.�.������� (1982:13-14) �����
���� ���������� � �������� �� �������������
����� ��������� (��� �������, ��� �� ���
���������� �� �������� �������������� �������
����� �������� �����, ��� ������� ��), �
����������� �� �� �.�.��������� �������
������������ ������������� ���������������
������.
� ����� ������ ���������� ����� ������ �����
���� ������ ������ ��������������. �������� �
������ � ������������ ����� �������� �����
��������� ��������������� �������� � ������� �
������������ �������������� ����������, �,
������, ����� � ������� ������� ������� ��.
������� ��������� ����� ����������� ������ ��
�������� ����������������� �������� �����
��������, �� ����������, ������ � ��������
���������, ������� ������ ��������� � �.�. ��� ���
���������� ���������� ��������� ��� ������� �
����� ��� � ������������ ������������� �������,
������� �������������� (�����, 1978) ���������
(�.�.����� � �� ������ ���������).
� ����� ������ �������� ��������������
������������ ������������� ������� �
����������� �������� ����� ����� ���� ������, �
������:
- ��������� ������� ��������� ���������
��������� ������� ������������� �������,
���������� ���������� � ����������������
��������� ������������� �����;
- ������ ������ ����� �������� � ������� ��������
��������� ����� ����������� ����������� ��
��������� ���� (�������� ��� ������������ ����- �
������������������ ��������������);
- ��������� ������� ����������-���������
�������� �� ��������������������� �����������
����� �������� � �� �������������
������������������ �������� �� ������� � ��� ���
���� �����������. ��� ����� ���������� �� ������
���������, �� ���������� ���������
������������� ��� ��� ���� ��������;
- ���� ������� ������������ ��������� �����
����������� ���������� �� ��������� � ���������,
������ �� ������ �������������, �� � �����������.
���������� ����� ������ ������� ��������������
�� ���������, �������� ������� � ������
����������� ����������� ������������, � �����
�������� ���������� �������������
(�����������������; �����������, 1981�) ����, �����
����� ������ ��������;
- �������� � ����� ��������� � ������� ���
������� ������� �������� �� ������� ������ �
���������� ���������� ����� (�� ������- �� ����- �
���������������);
- ��������� ������� ������� (������ �
����������������� ������������� ���������� �
������������� ���������) ����� ���������
�������� � ����� ������������� ��������� �����,
�������� ���� ��� ���� ����������� ��������. ���
�� ������� ����� ����������� ������ �� �����
��������� ����� � �������� � �� ���������
�������� ���������� (� ����� ����������
����������� ��� ��� ���� ��������� ������ ����
�����);
- ������������� ��������� ��������� �������� ��
������������ ����������� ���� �������������
������ (�������, 1995�)12.
����, �� (� ������������� �������) � �����
������ ������������� ����� ���������. ������
����������� ������������ ����� ���� �����������
������ �� ������. � ���� ������� �� �������������
����� ���� ��������� ����������� �����
������������� ��������� (��������, �������,
1969). ���������� ������ �������� �������� ������
��� ����������� ����� �-�� �������� (�����,
����������, 1994). �� ����� ��������������� � ����� �����
����������������� ����������� ��������� � �
��� ������, ���� �� ��� ���������� ��������
�������� ������������ �����.
��������, � ������ ������� ����������
����������� ���������� (� ��������� ��
������ �������� �������, ��� �������� �������
��� ������ ������� ����������� ����),
��������������� ������ ������� ��������
�������, ����� ���� ���� ���������� �����
������� � ���� ��. �������, �������� ����� �����
���������������� � ��� ������� ����������
����������� �� (� ������������ ������������
����� ������� ������������ ��������).
�������, ������� ������������� ������ �� ��
���������� ������������� ���������������
������, ������������� ��������� �������, ����
� �� ����������� ���� � ������ �����������
������ (�����, 1982, 1987�, 1987�; �����, �������, 1987�, 1991).
�������� ��������� ������������� ��������
������� �� �� �������� ���������������
����������� ������ ��� ���� �� ��������
�������������� (������������ ���������
����������-������������� ���������������
��������������; ��������, 1961; �������� � ��., 1994).
������ �������� ����� ��� ��������. ��������
������������� ������������ ������������� ����
�������� ��������� ����� ����� � ��������
�������� � ����� �� ������ �� ��������, �� ����
����������� �������� ������������ ��������
(��������� ������ ���������� ����� ����
������������ ���������� ��� ����������������;
��������, 1964; ���������, 1976). ���� �� ���������
������������ ������������ �������� ���
������������� ��, ����� �� �������
���������������� �������� � ���������� � �
��������� ����������� � ��� �����.
� ���������� ������� ���������� ���������� �
������ ��������� (���� � �� �������������)
�������������� ������. ���� ����������� �� �����
���������� ���������� ��������������������
������� �������� ������� �� ��������� �������
������������ ��������, �� ��������, ��� ��� ��
���������� �� ���� ����� ��������, � ��������
������������ ���������� ������. ��� ���� ������
���������� ����������� ��������� (���������� �
������� ���) ���������� �������� 105��: 10 -13��
� ������� ������� ����, 10-8 � ����� ������,
10-3 � ���� ������, 102 � ����������;
���������� ��, ����� ��� � ������������� �������
� ����� (� ����� �������� ������ � ����������
������), ���������������� �������� ����������� �
�������� ������� 10 7�� (�������, 1983). �����
�������, ��������� ������� ��������
������������� ��������� ������������� ������ �
�� � ����� ������ ����� ����������� �������
����������, ��� �� ������� ������ ���
�.�.������������, � ��������� ��� ������ �������
�������� ������� ��������� �� � � �����
����������� � ���������� �������������
��������� ���������� � �� ������������ ������.
����������
� ������� �� �����, ����������� �� �����
�������, ������������� ������� ������
��������������� ������� � �������, � ������
��������� ���������� ������������� ����� ��
�������. ��������� ���� � ������ ������ �
������������ ��������� ����� ���������������
��������� �� ���������� ����������
�������������� ������������, ���������� ��� �
������ ��������� ��� ������� ��������������. ���
������ ���������� ������ �� � ��������������
�������� � ���� ��������� �������������� ������
�������������. �������� ����������� � ����� �
���������� ������ � ��� ������� ������ ����
��������� ����� �� �������� ����������.
������������ ������ ������ �� �������� ������ ��
� �������������� ����, ��, ��� ������� ������,
������������ �������� � ���� ����������
(���������� � ���������� ��������� �����).
������ �������, ����� ������ ���, ����������
�.�.�����������, ������ ������� �������������
���� ��������� ������, ������������ �� �� ������
������� �������, �������������� � ��� �����
��������� ������������� ���� (��������, 1992).
��������, ������ ��������� �� �������� ���������
��� ����� �������� ������ �������������
������������ ���� (�����, �������, 1987�, 1991), ����
������ ����� �������� �����.
����� �������� ����������� �.�.������,
�.�.��������, �.�.���������, �.�.���������,
�.�.���������, �.�.���������, �.�.���������,
�.�.���������, �.�.������� �� ������ ������ ���
��������� ������, �.�. �������� � �� ������ ���
���������� ������������, �.�.��������,
�.�.��������, �.�.���������, �.�.�������� �
�.�.��������� � �� ������ ��� ���������� �������
������������ 1993 �.
����������
- ������� �.�. �������������� �������� �����
����� ������������ ����������� // ��������������
������������ � ������������ �����. �.: ����
��������� �����, 1981. �.5-19.
- ������������ �.�. �������� ����
������������� ������� // ������� �����������. �.;
�.: �����, 1964. �.3. �. 300-447.
- �������� �.�. �������������� ���������� ��
�������� ������� ������������� ������������� //
���. ����, 1978. �.83, � 2. �.138-142.
- �������� �.�., ��������.�. ����� ������������
������-��-���. �.:���-�� ���, 1980. 104 �.
- �������� �.�., ��������� �.�., ���������� �.�.
������������ ������ ������������ ������. ������:
���� ���, 1992. 177 �.
- ������ �.�. Anemone nemorosa L. � ����������
�������� // ��. �-�� �������� ����. ���. 1930. ���.4.
�.51-75.
- ������� �.�., ������������� �.�., ������� �.�.
����� ������ �������� ����������� ���
(���������� � ������������). ���.: �����, 1991. 206 �.
- ��������� �.�., ����� �.�. ����� ����������
�������� ����-�������� ���������� // ��.
�������������� ���. �����������. ��������:
��.���-��, 1969�. ���.7: ������������ ������������.
�.3-59.
- ��������� �.�., ����� �.�. ����� ����������
�������� ������� �������� // ��. ��������������
���. �����������. ��������: ��.���-��, 1969�. ���.7:
������������ ������������. �.126-177.
- �������� �.�. �������������� �������������
������-������ � ������ ����������� ����� ������:
������� � ������: �������. ���. ... ����. ����. ����.
���.: �����, 1992. 17 �.
- ������� �., ������ �.�. ������ ������������
���������. �.; �.: ���������, 1936. 715 �.
- ��������� �.�. ������ �������������
�������������. �.: �����, 1983. 248 c.
- ������� �.�. ��������������� ���������
�������� // ���������. ����. 1987. � 1. �.3-12.
- ������� �.�. �������������� �������������
��������� ����������� // ������ �����������
��������. 1993. �.113, ���.5. �.564-575.
- ��������� �.�. ���� ��������������
������������ �� ������ ��������� �����������
����� (�� ������� ���������� �����������) //
������������� � ������������ ��������
������������� ����������. �.: �����, 1987. �.90-104.
- ��������� �.�. �������� ������������
��������� ������ ��������� // ���� ����� ������.
�.: ���-�� �� ����, 1963. �.5-29.
- ������� �.�. ������� �������� �������� �
������� � ��������� �������� ���������� //
������������� � ���������������� ��������
������������� ����������. �.: �����, 1987. �.36-42.
- ������� �.�. ������������� (�������� � ����
��������) // ���������� �������� ��������������
�������� ����. ���.: �����, 1994. �.116-132.
- �������� �.�. ���� ���������������
������������� ���������� ������� // ���. ����. 1964.
�.59, � 4. �.142-144.
- �������� �.�. �������������� // ��������
����� ��������� ������. �.: ����. ����-���, 1975.
�.12-29.
- �������� �.�.�������������� // ������������
�������� ������. �.:���-�� ���, 1986. �.1. �.16-32.
- �������� �.�., ������� �.�., ��������� �.�. �
�������� ��������� ���� (�� ������� �������
������������) // ���������� ��������
�������������� �������� ����. ���.: �����, 1994.
�.87-96.
- ������ �.�. ������� ����- � ���������� ��
������� ��������� ������������� �������� //
�����. ����. 1965. �.50, � 8. �.1121-1126.
- ������� �.�. ���������� � ��������� � ������
��������� ����� ������������ ����������� �����
(��������� �-��) // �����.����. 1993. �.78, � 2. �.126-134.
- ������� �.�. ������������� �������
�������������� � ������ �� ��������. ��������
�������������� ������� ���� // �������������
�������� ������������� ������� ������� �
����������: �������� � ������ ��������. ���.: ���
���, 1995�. �.115-135.
- ������� �.�.��������������� ���������
������������ �������� // �������� ���������: ���.
����. �����������. ������., 26-30 ����. 1995�. �.: ���
���, 1995�. �.69.
- ������� �.�. ���������� ��������
�������������� �������������� ������� (��
������� �������� �.�������): �������.���. ... ����.
����. ����. ���.: ��� ���, 1996. 26 �.
- ��������� �.�. �������, ������������
�������� ������������� // ���. ���. 1976. �.99. �.75-80.
- �������� �.�., ������� �.�. ���������� ����� �
����� ��������� // �����.����. 1969. �.54, � 12. �.1911-1920.
- ����� �.�. �������� ������� ���������
���������� // ����.����� ��������. 1978. �.39, � 4.
�.495-508.
- ������� �.�. �����������. �.: ����. ����-���,
1980. 408 �.
- ������ �.�., �������� �.�. �����
����������-������� ����������������
�����������. �.: �����, 1976. 104 �.
- ��������� �.�. ������� ������ �����
��������� � ������ �������� ������������
����������� // ��. ������������ ���. �����������.
�.: ��������� �����, 1960. ���.4. �.127-188.
- ������� �.�., ������� �.�. ����������
�������� ����������� �����������. �.: ������, 1987.
52 �.
- ������� �.�. � ������������� // �����. ����.
1965. �.50, � 6. �.797-810.
- ������ �.�. � ���� ������������� �
���������������� �������� � ��������������� Anemone
nemorosa L. // �������-������������ � ��������������
����������� ��������������. �.: �����, 1983. �.173-184.
- ������������ �������� ������. �.:���-�� ���,
1986. �.1. 240 �.; �.:���-�� ���, 1987. �.2. 224 �.
- ����������� �.�. � �������� ������� �
�������������� ������ // ���. ���. 1969. �.72. �.20-28.
- ����������� �.�. ��������-��������������
������������� ����� ��� ����������� ��������
����������� �������� // ����������� �����������
� �������������� ��������. �.:�����, 1981�. �.10-27.
- ����������� �.�. �������������� ��������
����������. �.: �����, 1981�. 232 �.
- ����������� �.�.� ��������� ������ ��������
� ������� ����� ���� � ������������ �����������
// �����.����. 1997. � 3. �.1-6.
- ����������� �.�. �������� � ���������
������������ ������������� �������. �������� �
�������������� ������������� �������. ��������
������ ��������-��������������� �������������
�����. �.: KMK Scientific Press, 1999. C.17-236.
- ��������� �.�. �������� � �����������
��������-��������������� ������������ ������.
�.: ����������, 1938. 620 �.
- ��������� �. �. ������ ����� ����������
������� � �������. �.:�����, 1983. 216 �.
- �������������� � ����� ��������������
������������� ������� ���. �.: ���-�� ���, 1986. 90 �.
- ��������� �.�. ���� ���������� ������
���������� ���� � ���������������� �������
(���������� ����) // ������������� � ������������
�������� ������������� ����������. �.: �����, 1987.
�.67-90.
- ����������� �.�. ��������� ����������
������������� �� �� �������������� ���������
(������ ���������� �����������) // �����. ����. 1989.
�.74, �4. �.498-508.
- ��������� �.�. ����������������� ��� �����
������������ ������ � ������������ �������
���������� �����. �.: �����, 1981. 232 �.
- �������� �.�. �������������� ��������� �
����� ����������-������� ����������������
����������� (����������� �������) // �����. ���.
���.����. 1980. � 3, ���.1. �.67-70.
- ������� �.�. ����� ��������-����������
���������������� ����������� //
��.��������-���������� ���.�����������. �.:
��������� �����, 1958. ���.2. 248 �.
- ������� �.�. ���������� � ��������� �
���������� ����� // ���.����. 1971. �.76. ���.4. �.128-133.
- �������� �.�. ��������� ��������� �������
���������������� �����. �.: �����, 1987. 205 �.
- ������ �.�. ������������ ����� �����-����� //
�����. ����. 1932. �.17, �2. �.185-202.
- ������� �.�. ��������� ����� �������������
������� ������������� // ������� ��������. �.; �.:
���-�� �� ����, 1954. �.1. �.291-309.
- ������� �.�. �������� ���������� ��������� �
������ ������������ ������ // �������� �������
������������� ������. �.: ���. ���, 1983. �.90-112.
- ������ �.�. ����� ������ � ����� ����-������
// �������-�������������� ������������ � ����� �
������� ������� � ���������. �������: ������. ���.
�� ����, 1980. �.83-86.
- ������ �.�. ��������� ����� ��������������
�������� ����� ���������
��������-��������������� ������������� ������ //
������������� � ������������ ��������
������������� ����������. �.: �����, 1987. �.104?107.
- ��������� �.�., �������� �.�., ��������� �.�.
�������������� ������ ���������� ��������
������� �����������. �.: ������, 1987. 78 �.
- �������� �.�. � ��������
������������-�������������� ������������.
������� � ����� � ������������� ���������� //
����. ���. 1931. �.16, �1. �.111-124.
- �������� �.�. ������ ������ �� �������. �.:
���-�� ���, 1962. 100 �.
- �������� �.�. ��������� ���� ��� ������
�������������� �������� // �����.���. 1970. � 9. �.71-83.
- �������� �.�. �������� � ��������� ��������.
�.: ���-�� ���, 1974. 244 �.
- ������ �.�.� �������� ���������� ����
������� ���� Piceetum sorbosum (������ ���������) // ���.
��������. 1939. � 1. �.107-113.
- �������� �.�.����� ����������� �������. �.:
�����, 1985. 398 �.
- ����� �.������ ����� ��������� ��������. �.:
���-�� �����.���., 1956. 380 �.
- �������� �.�. �������� � �����������. �.:
���-�� ���, 1964. 447 c.
- ����� �.�.����� ������-�����: ��������
������� ������������ ���������� ������-�������
������. �.: �����, 1968. 234 �.
- ����� �.�.�������� ������������ ���������
������-��������� ����. �.: �����, 1974. 160 �.
- ����� �.�.��������� ��������� ��������
������ ���������� ���� // �����. ����. 1975. �.60, � 1.
�.69-83.
- ����� �.�. ����� ��� ��������� ������� // ���.
����. ���.����. 1982. �.87, ���. 4. �.3-22.
- ����� �.�.��������� ������� ��������
��������� �������� � �������� ��������
��������� ������� // ��������� ���������� �
�������� �������� ������: ����. ��������. �����.,
�������, 27-30 ����. 1984 �. �������: ���-�� �� ����, 1984.
�.207-212.
- ����� �.�.������������� ��������� �
��������� ����� �������� // ��������� ����� �
�������� � ����������� ��������: ������. ��.
����.��. �.: ���-�� ����, 1986. �.9-23.
- ����� �.�. ����� ��� ������� �������
����������: ���������� �������, ������� �
�������� // ������������� � ������������
�������� ������������� ����������. �.: �����. 1987�.
�.13-28.
- ����� �.�.������������ ������������ ����� �
������� ������� ������������� ���������� //
������������� � ������������ ��������
������������� ����������. �.: �����, 1987�. �.47-66.
- ����� �.�.�������� ��������� ����������
���� �������������� // �����. ����. 1991. �.76, � 1.
�.30-41.
- ����� �.�.�������-�������������� ���������
�������������� ������������ � ��������� ���
����� � ������ // ������������� ������������:
������� � �������� � ����������. ���.: ��� ���,
1992�. �. 7-21.
- ����� �.�.��������� �������� � ���������
������������ ���� �������������� (III-IV 1992,
�����-���������) // �����. ����. 1992�. �.77, �10. �.102-106.
- ����� �.�. � ��������� �������������
�������� ������������� ���������� // ����������
�������� �������������� �������� ����. ���.:
�����, 1994. �.15-33.
- ����� �.�. ������ ������ ������������������
���������� ����� // ������������� ��������
������������� ������� ������� � ����������:
�������� � ������ ��������. ���.: ��� ���, 1995.
�.136-146.
- ����� �.�.,������� �.�.���������
�������������� ������������ ������ �������
����������� // ������������� � ������������
�������� ������������� ����������. �.: �����, 1987�.
�.219-242.
- ����� �.�.,������� �.�.����� ������� ��������
������� ���������� // ������������� �
������������ �������� ������������� ����������.
�.: �����. 1987�. �.242-266.
- ����� �.�.,������� �.�.�������� ������� �
������� ����������. �����: ���-�� ����. ��-��, 1991. 80
�.
- ����� �.�., ������� �.�.������ �������������
���� ��������� ��������� ��������� �������
������ (����� ���������� �������) // �����. ����.
1993. �.78, � 3. �.11-32.
- ����� �.�.,���������� �.�. ����� ������������
����� ������������: ���������� �������� //
����������� ������ ������� ��������. ���.: ���
���, 1994. �.7-66.
- ����� �.�., ���������� �.�., �������� �.�., ��������
�.�., �������� �.�. ����� ���������������
��������������� ���������� �������� ���������
������. �����.1 // ���.����. ���.����. 1979�. �.84, ���.5.
�.111-122.; �����.2 // ���.����. ���.����. 1979�. �.84, ���.6.
�.74?83.
- �������� �.�.�����������. �.; �.: ���-�� ��
����, 1961. 474 �.
- Bagyaraj J. Ecology of vesicular-arbuscular mycorrhizae // Handb. Applied
Mycology. N.Y.: M.Dekker Inc., 1991. Vol.1. Soil and Plants. P.3-33.
- Braun-Blanquet J., Pavillard J. Vocabulaire de sociologie vegetale. 3 ed.
Montpellier: impr. Roumegous et Dehan, 1928. 23 p.
- Dansereau P. Biogeography: An Ecological Perspective. N.Y.: Ronald Press, 1957.
394 p.
- Grime J.P. Plant strategies and vegetation processes. N.Y.: Wiley, 1979. 222p.
- Harley J.L., Smith S.E. Mycorrhizal symbiosis. L.; N.Y.: Acad. Press. 1984. 483p.
- Sumina O.I. Plant communities on anthropogenically disturbed sites of the
Chukotka Peninsula, Russia // J. Vegetation Science. 1994. Vol.5. P.885-896.
- Tansley A.G. The use and abuse of vegetational concepts and terms // Ecology.
1935. Vol.16, N 3. P.284-307.
- Van Valen L.The Red Queen // Amer.Natur. 1977. Vol.111, N 980. P.809?810.
- Watt A.S.Pattern and process in the plant community // J.Ecol. 1947. Vol.35.
P.1-22.
- Watt A.S. Bracken versus heather: A study in plant sociology // J.Ecol.
1955. Vol.43. P.490-506.
1 �������� �� IV ������� ��������� ��
������������� ���������� (�����������
���������� ����������, 17-20 ����. 1993�.)
2 ������������ ���������� ��������
���������� �� ������ �.�.������ � ��. (1979�,1979�) ���
�������, �� �.�.��������� (1982) ��� ������������ �
�� ������������� �������� ������� (1986, 1987) ���
������� ������
3 ����������� �� ���������� ����� ��
��������������� ������������� �������
������������� ������, ��.��������������, 16
������� 1980 �. ������������� �� ������������
������������� ������
4 ��� ���� ���������� � ���������������
�� ������������ ���������� ����� � �� 70-90% ��
�������������� ������ ��� ������� ���
(�����������, 1981�). ����������� 10-30% �������������
�� ������� ����������� �����-���� �������������
��� (� ��������� ���� ������������ ����), �������
���������� ������ �� ������������� ����������
�����, ��������� ��� �������� ����, �����������
�.�.����������� sensu lato, ���������� ��������� �
������������ � ����� ����� �� ���������� �
�������������� ������� ��
5 ��������, ��������� �������
����������� � � ��������� ���������������
����� � ���������, ������������� �
��������������. ���, ������� (���� ���� ��� ��
������������ �������) ��������������� ��������
�������������� � ���� ��������������� ���������
���� ����� ����� Salicaceae � ���� ��������� �� ������
�������� �� ������-������� ����. � �����, ������,
����������������� ��������������� �����������
����. ��������, � ����������� �������
����������� ��������� ���� �� �����
��������� �������� �������� ��� �������
�������������� ��������� �����. ������, �����
�������������� �������� �������������� �����
���������� ���� ����� ������������ ������.
��������, �������� ����� �������� ���������� ��
������ ��� � �� ������� �����, ���, ��������,
������������ ��������� ���� ���������������
���������.
6 ����������� ����� Carex �����������
��������� ��������������; ������, ��� �������
�������� ������������� � ������� �����������
���� � ������ ��������������� ����������.
7 ����������� �����, ������, �������� �
����������� �� �������� ����: ������� �������,
����������� Dryopteris carthusiana, Paris quadrifolia � Rubus saxatilis
8 ��������������� ��������� A.glutinosa,
��������, ���� ��������� ������� A.incana, ������ �
��������� ������������� ���� �������� ��������
����������� (���������) ������������ ���������
9 ����� ������ ������ � ������������
���� ����� ������ Nanae, ������� �.�.����� (1986)
�������� ����������� ��������������� �������
���������� �� ������ �����������; ��������, ���
��� ���� � ������������� ����������� � �������
������ �����. � �� ������� ���� ��� ����������� �
������������� � Dryas punctata, ���������� �����������
����� ������� ��� ��� ������ ���������, ��� �
����� ����, ��� �������� � �������������
���������� ������ (�������, 1996)
10 �.�.����������� (1981 �, �.160) ������� ����
� ���� �����, �������� � ����������� �������
������������, �.�. � ��������� � �������
������������� ����������� �����
11 �������, ������� � ���� �� �������
���������� ������, � ������ �� ��������, �������,
������, �������� � ����� �� ���������������
�����
12 � ����������� ���� ���������� ���
������ ������������� ������, ������ ��� ���� ���
������ �� ��������������� � � ������ �������,
��-�� �������������, ������������� �����
������������� ������������ ����� �
���������������. ������� ������� ���������
��������� �� �����������, � �� ������
�������������, �� ���� ��� ��� �������� ���
������ �������
***
������ ������ ���� ������������ � ������� �������������� ������ ������������� �����
"������� ����"
������ ����������������� ������, ��������� � ������������� ������ ����, ����� ����������
� ������� ����������������� ������. ������ ������� ����������������� ������ (����������������� �������), ����������� �� ����� �����, ����� ����������
���.
 ������� ������
"�������� ����"
��������� "Parus"
������ ���������
������� ������
"�������� ����"
��������� "Parus"
������ ���������
|

